적요
Bakanae (foolish seedling) disease caused by Gibberella fujikuroi creates serious problems in the foremost rice growing countries. This study was conducted to identify new resistance genetic sources to Bakanae disease. Bioassay showed that 11 varieties including Gwangmyeongbyeo, Hawn, Wonseadaesoo, Erguailai etc. were resistant to bakanae disease among 254 rice germplasm. Mismatch ratio between phenotype on bakanae disease bioassay and allele type of RM9, a SSR marker closely linked the bakanae disease resistant QTL, qBK1, were 38.3%. These results suggest that RM9 might be used for selecting qBK1, but it cannot be used for wide range of rice germplasm. Resistant germplasm in this study might be have resistant genes different from qBK1. The eleven varieties resistant to selected in this study will be used to identify new resistant alleles or genes to improve bakanae disease resistance in rice.
서 언
벼 키다리병은
Gibberella fujikuroi에 의해 발생되며 일본 에서 처음 보고되었다 (
Hori 1898). 본 병은 아시아 지역에서 주로 발병되나 최근 세계적으로 상자육묘와 기계이앙의 발달 과 하이브리드 벼 재배 증가와 함께 발병이 확산되는 추세이 다(
Rood 2004,
Rosales & Mew 1997,
Yang et al. 2003). 미국에서도 1999년 본 병이 최초로 발견된 후 캘리포니아 지 역 대부분의 벼 재배지역에 확산되고 있다(
Carter et al. 2008). 우리나라에서는 2000년대 초반 까지는 전국 벼 재배지의 약 3% 정도에서 키다리병이 발생되었으나, 2006년에는 전국 논 의 28.8%(
Han 2007), 2013년에는 31%에서 키다리병이 발 생이 확인되었다. 유묘에서 벼 키다리병의 일반적인 증상은 비정상적인 초장의 증가와 함께 줄기는 가늘어지고 식물체의 색깔이 황록색으로 변하는 것이다.
Yamanaka & Honkura (1978)는 키다리병 증상을 (a) 신장, (b) 신장 후 정상생육, (c) 신장 후 초장단축, (d) 초장단축, (e) 생육중단 또는 고사 등의 5가지로 분류하였다. 이러한 증상은 병원균에 의해 생성 되는 지베렐린과 fusaric acid의 비율에 의해 조절된다(
Ou 1985). 성숙기 벼에서도 감염식물체는 절간장의 신장 또는 단 축이 확인되며 줄기가 얇고 하위 엽부터 갈변되어 심한 경우 식물체가 고사하거나 생존하더라도 등숙이 매우 불량하게 된 다(
Mew & Gonzales 2002). 벼 키다리병에 의한 수량손실은 일본에서는 지역에 따라 20 ~ 50%(
Ou 1985), 1993년 인도의 경우에는 3.04 ~ 96.25%까지 감소한 것으로 보고된 바 있다 (
Sunder et al. 1997).
벼 키다리병 저항성 품종을 선발하기 위한 기존의 연구에서 접종원으로는 병원균의 포자(
Iqbal et al. 2011,
Khokhar & Jaffrey 2002,
Lee et al. 2011,
Ma et al. 2008,
Park et al. 2009,
Sukapanpotharan & Chantarasnit 1980,
Yang et al. 2003,
Zainudin et al. 2008), 자연 감염된 토양(
Saremi et al. 2008) 등이 이용되었으며 검정환경으로는 포장(
Iqbal et al. 2011,
Khokhar & Jaffrey 2002,
Saremi et al. 2008), 온실 (
Lee et al. 2011,
Zainudin et al. 2008) 및 생장상(
Park et al. 2009,
Yang et al. 2003,
Sukapanpotharan & Chantarasnit 1980) 등 다양한 환경에서 검정되었으나 시험 시기 등 환경에 따라 서로 다른 결과들이 도출되었고, 포장저항성 검정의 경 우 많은 시간과 공간이 요구되는 문제점으로 대량의 품종을 검정하는데 그 실효성이 낮은 단점이 있었다. 이러한 단점을 개선하고 대량의 자원을 동시에 검정하기 위해
Kim et al. (2014)은 Tissue embedding cassette
TM를 이용하여 신속하고 균일한 접종법을 확립하고 키다리 병원균의 포자농도, 접종 후 저항성 평가기준 등을 설정하여 저항성 대량 검정체계를 구축한 바 있다.
저항성 품종을 육성하고자 할 때 전통적인 육종 방법은 유 용 유전자원 도입을 위해서 대량의 유전자원 또는 계통을 생 물검정을 통해 저항성 여부를 판단하여야 하므로 저항성 품 종을 개발하기까지는 오랜 시간과 노력이 동반되어야 한다. 또한 저항성 검정에 있어서 온도, 광량, 습도, 식물체의 생육 정도 등 각종 검정조건에 따라 저항성의 정도가 달라 저항성 품종을 선발하는 데에 어려움이 있어 가장 효율적인 검정방 법인 분자표지방법이 매우 필요한 실정이다. 벼 키다리병에 대한 유전연구는 세계적으로 매우 부족한 실정이다. 중국에서 는 Chunjiang 06/TN1 DH 집단을 이용하여 1번 염색체와 10 번 염색체에서 저항성 QTL을 확인한 바 있으며(
Yang et al. 2006), 우리나라에서는 일품/신광 BC
6F
3 집단을 이용하여 1 번 염색체에 존재하는 신광벼 유래의 저항성 QTL인
qBK1을 확인하고 연관마커인 RM9를 선발하여 보고한 바 있다(
Hur et al. 2015).
본 연구는 벼 키다리병 저항성 유전자원 다양화를 위해 유 전자원에 대한 저항성을 평가하고 신광벼에서 유래한 qBK1 영역의 유전적 다양성을 분석하여 새로운 저항성 자원을 확 보하고자 수행하였다.
재료 및 방법
실험재료
국내외에서 육성 또는 도입된 품종 및 유전자원의 키다리 병 저항성 검정을 위해 국내 자포니카 141종과 통일형 26종 및 국외도입 자포니카 64종, 열대 자포니카 3종, 인디카 20종 을 포함하는 총 254종의 유전자원을 이용하여 본 연구를 수 행하였다.
키다리병 저항성 검정
벼 키다리병 저항성 검정은
Kim et al. (2014)의 방법에 준 하여 수행하였다. 농촌진흥청 국립농업과학원으로부터 분양 받은
G. fujikuroi CF283의 균사를 potato dextrose agar (PDA) 배지에 치상한 후 28°C 명 조건에서 3주간 배양하였 다. 배양된 균사체는 증류수에 현탁 후 포자농도를 1×10
6 spore/ml로 희석하여 접종에 이용하였다. 균 접종 직전 품종 당 30립의 종자를 Tissue Embedding Cassette(Simport, M512, 40.1×28.5×13 mm)에 넣고 57°C에서 13분간 온탕 처리 하여 종자소독을 실시하였다. 온탕처리된 종자는 찬물에 서 열을 식힌 후 준비된 포자배양액에 침지한 후 26°C에서 4 일간 접종하고 매일 4회 이상 Tissue Embedding Cassette를 천천히 돌려 포자가 모든 종자에 고르게 접종되도록 유도하 였다. 접종된 종자는 육묘용 상자에 상자당 20품종씩 배치하 고 품종당 30립의 종자를 파종하였다. 처리의 균일성을 확인 하기 위해 비교 품종인 주남벼와 남평벼를 파종하여 상대적 인 저항성정도의 기준으로 이용하였다. 육묘용 상자에서 발아 된 종자는 온실에서 생육시킨 후 검정에 이용하였다. 저항성 판단은
Kim et al. (2014)의 방법을 응용하여 이앙 1달 후 품 종별로 총 식물체 수에 대한 이병주의 비율과 육안에 의한 저 항성 정도 0(저항성) ~ 9(감수성)로 구분하여 판별하였다.
Genomic DNA를 추출하기 위하여 어린 벼 잎 3 ~ 4장을 2 mL tube에 3 mm 텅스텐 구슬 3개와 함께 넣고 액체질소 에 5 ~ 10분간 침적시킨 다음 vortex하여 마쇄하였다. 마쇄된 샘플은
Chen & Ronald(1999)의 방법에 따라 cetyltrimethyl ammonium bromide(CTAB)를 사용하여 genomic DNA를 분리한 후 PCR 분석에 이용하였다. 품종별 키다리병 저항성 유전자형 분석에 이용된 primer는 통일형 벼인 신광벼에서 유래한 벼 키다리병 저항성 QTL인
qBK1를 표지하는 RM9(23.39 Mbp)를 이용하였다(
Hur et al. 2015). PCR 조건 은 denaturation을 94°C에서 30초, 55°C에서 30초, 72°C에서 60초로 40 cycle을 실시하였다. PCR 반응을 끝낸 증폭 산물 은 3% agarose gel에서 전기영동을 실시하여 EtBr로 염색한 후 genotype을 조사하였다.
결과 및 고찰
벼 주요 유전자원의 저항성 검정 결과
환경변이 등에 따른 키다리병 저항성 판정의 신뢰도를 높 이기 위하여 국내품종 중 저항성 정도가 비교적 높은 것으로 알려진 남평벼와 감수성 품종인 주남벼를 매 10품종 마다 파 종하여 총 26반복에 대한 이병주 비율의 분산분석을 통해 전 체 시험품종의 단반복 저항성 성적에 대한 정확도를 유추하 고자 하였다. 비교품종의 반복력에 대한 분산분석 결과, 품종 간에는 고도의 유의성이 인정되었으며 반복간에는 유의성이 없었다(Table
1). 따라서 본 연구에 이용된 공시품종에 대한 저항성 검정성적의 정확도는 인정될 수 있는 것으로 판단되 었다.
Table 1.Analysis of variance for proportion of healthy plant on ‘Junam’ and ‘Nampyeong’ on rice bakanae disease bioassay
Table 1.
|
Source of variance |
DF |
Sum of square |
Mean square |
F Value |
|
|
Variety |
1 |
35485.6 |
35485.6 |
177.59z
|
|
Replication |
25 |
5141.6 |
205.6 |
1.03ns
|
|
Error |
25 |
4995.1 |
199.8 |
|
|
Total |
51 |
45622.6 |
|
|
본 연구에서 저항성 생물검정 결과 키다리병에 대한 완전 한 저항성 품종은 확인되지 않았으나, 기존 연구(
Kim et al. 2014)에서 저항성으로 분류된 신광벼 수준의 저항성을 가진 품종군을 중도 이상의 저항성(저항성 정도 4이하, 이병주 비 율이 30% 미만)으로 분류하고, 감수성 품종인 주남벼 수준인 품종군을 감수성(저항성 정도 7이하, 이병주 비율이 70% 이 하)으로 분류하였으며 그 사이에 있는 품종을 중도 저항성 및 중도 감수성으로 분류하였다.
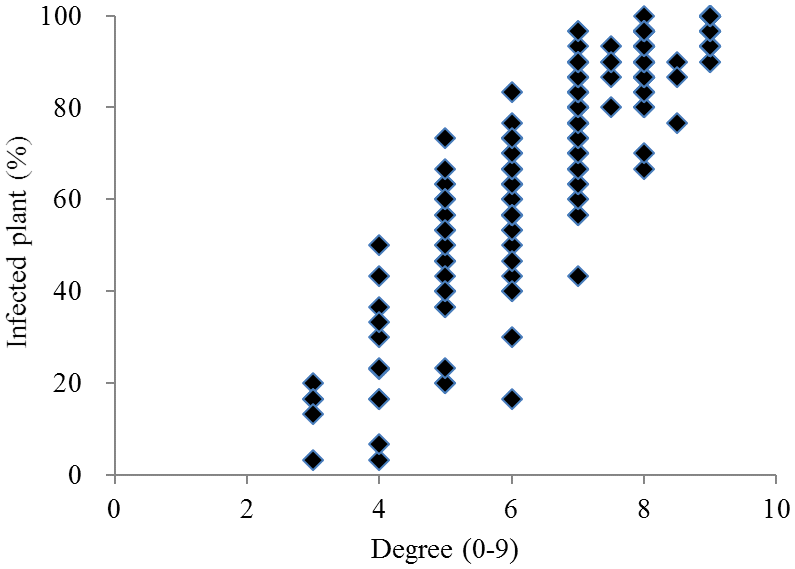
국내외에서 육성 품종 및 도입 유전자원 254 품종에 대한 키다리병 저항성 검정결과(Table
2, Fig.
1, Fig.
2), 중도 이 상의 저항성 품종은 11품종으로 전체 공시품종의 약 4%정도 이었으며, 인디카형에서는 저항성 정도가 높은 품종은 확인되 지 않았다. 국내 육성 자포니카 품종에서는 광명벼(저항성 정 도3, 이병주 비율 20%), 통일형 품종 중에는 신광벼(저항성 정도4, 이병주 비율 23%)가 중도 이상의 저항성을 보였다. 또한 새누리, 동진벼 등 국내육성 자포니카 벼 9품종은 저항 성 정도가 4 ~ 6이며 이병주 비율이 50% 미만으로 나타났으 며 나머지 131 품종은 저항성 정도가 낮거나 매우 약한 것으 로 나타났다.
Table 2.Classification of resistance to bakanae disease on 254 rice varieties
Table 2.
|
Origin |
Type |
Number of varietyz
|
|
|
Over medium resistance (≦4z, <30%y) |
Medium (4~6, ≦50%) |
Medium susceptible (4~6, ≦70%) |
Susceptible (7≦, 70%≦) |
Total |
|
|
Korea |
Japonica
|
1 |
9 |
18 |
113 |
141 |
|
|
Gwangmyeongbyeo |
Aranghyangchalbyeo |
Danmi, |
Boramchan |
|
|
|
|
Cheongnam, Goun |
Dongjin 1 |
Dongjin 2 |
|
|
|
|
Hwananmbyeo |
Saesangjubyeo |
Geurubyeo |
|
|
|
|
Dongjinbyeo |
Hwanggeumbora |
Gopumbyeo |
|
|
|
|
Saenuri, Boseog |
Chilbo, Goamibyeo |
Hopum, Ilpumbyeo |
|
|
|
|
Baegjinju 1 |
Dongjinchalbyeo |
Junambyeo |
|
|
|
|
Mangeumbyeo |
Malgeumi |
Junamjosaeng |
|
|
|
|
|
Samgwang etc. |
Seolgaeng etc. |
|
|
|
|
Tongil
|
1 |
3 |
5 |
17 |
26 |
|
Shingwangbyeo |
Areumbyeo |
Dasan 1 |
Nogyang |
|
|
|
Geumgangbyeo |
Dasan2 |
Andabyeo |
|
|
|
Saegyejinmi |
Hanareumbyeo |
Cheongcheongbyeo |
|
|
|
|
Mokwoo |
Dasanbyeo |
|
|
|
|
Yongmunbyeo |
Hanareum 2 |
|
|
|
|
|
Hangangchal 1 |
|
|
|
|
|
Hangangchalbyeo |
|
|
|
|
|
Keunseom etc. |
|
|
|
Intro-duced |
Japonica
|
8 |
6 |
4 |
46 |
64 |
|
|
Bukruksa209 |
Chucheongbyeo |
Midorinomochi |
Akidagomachi |
|
|
|
Erguailai |
Dohoku 144 |
Princess Sari |
Aya |
|
|
|
Misamgeum |
Hongeui |
Yumechukusi |
Hinohikari |
|
|
|
NEP HUONG |
Joja, |
Yunhwae 422 |
Hitomebore |
|
|
|
Norin 1 |
Mochiminori |
|
IRAT13 |
|
|
|
Padi Adongdumarat |
Nipponbare |
|
Kirara397 |
|
|
|
Seonamna141 |
|
|
Milky Queen |
|
|
|
Wonseadaesoo |
|
|
Satojiman etc. |
|
|
|
|
|
Tropical
|
1 |
0 |
1 |
1 |
3 |
|
japonica
|
Hawn |
|
Pokharili-Mashino |
Kahei |
|
|
|
|
|
Indica
|
0 |
1 |
10 |
9 |
20 |
|
|
|
TW16 |
Apo |
Basmati 389 |
|
|
|
|
|
Basmati wx |
Dharial |
|
|
|
|
|
Chen Ma Ai |
Gumei 4 |
|
|
|
|
|
Congshengla |
IR64 |
|
|
|
|
|
Gumei 2 etc. |
IR72 etc. |
|
|
|
Total |
11 (4%) |
19 (7%) |
38 (15%) |
186 (73%) |
254 (100%) |
Fig. 1.Frequency distribution of the proportion of infected plants and degree of resistance (n = 254).
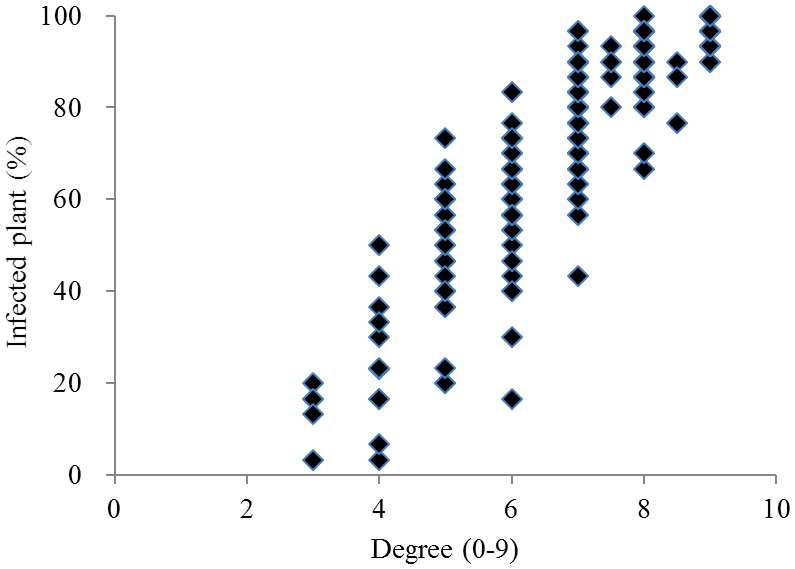
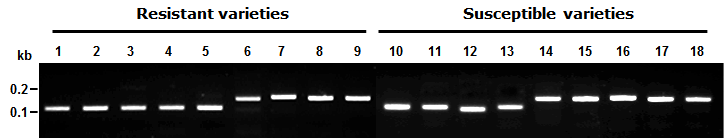
Fig. 2.Symptom of resistant and susceptible varieties on bakanae disease infection.

도입 유전자원 중에서는 중도 이상의 저항성 품종은 9품종 으로 국내 육성품종에 비해 도입품종에서 저항성 품종이 더 많은 경향이었다. 국내품종의 키다리병 저항성 품종의 출현빈 도가 외국 품종에 비해 상대적으로 낮은 이유는 육종과정에서 본 병에 대한 종자소독 등 방제를 통해 최근 발생되고 있는 키 다리병에 대한 자연선발이 되지 않는 조건으로 육종되었으며 또한 국내 육성 품종간에 유전적 다양성이 낮기 때문인 것으 로 추측된다. 도입유전자원 중 자포니카형인 Wonseadaesoo (저항성 정도 3, 이병주 비율 3%)와 Padi Adongdumarat(저 항성 정도 3, 이병주 비율 3%) 등 8품종과 열대 자포니카형인 Hawn(저항성 정도 3, 이병주 비율 13%)은 향후 정밀검정 후 각각 자포니카 및 통일형 벼의 저항성 품종 개발에 유용한 자 원으로 사용될 수 있을 것으로 판단된다.
분자마커 RM9를 이용한 저항성 관련 유전적 다양성 분석
신광벼에서 유래한 벼 키다리병 저항성 유전자
qBK1에 대 한 분자마커인 RM9(
Hur et al. 2015)를 이용한 벼 품종별 대 립유전자형과 벼 키다리병 생물검정을 통한 저항성 검정결과 와의 불일치성을 비교하였다. 분석에 이용한 품종은 분석의 신뢰도를 높이기 위해 본 연구에서 공시한 품종 중 중도 이상 의 저항성으로 구분된 11품종과 또는 감수성으로 확인된 품 종 중 156 품종을 대상으로 하였다(Table
2). 생물검정에서 저항성으로 구분된 품종 중 광명벼, Nep Houng, 북룩사209 호, 미산금은 RM9의 대립유전자형이 저항성 품종인 신광벼 와 동일한 유전자형을 나타낸 반면, Hawn, 서남나141호, 원 씨대수, Erguailai 등은 저항성 표현형을 나타냄에도 불구하 고 RM9에 대한 대립유전자형은 감수성 품종인 일품벼와 동 일한 유전자형으로 나타났다(Fig.
3). 총 167품종에 대한 키 다리병 저항성 검정결과와 분자마커 RM9에 대한 유전형과의 불일치성은 38%로 나타났다(Table
3). 이는
Kwon et al. (2012)의 연구에서 벼 유전자원 155품종에 대한 벼줄무늬잎 마름병 저항성 생물검정 결과와 저항성 분자마커에 InDel7에 대한 불일치성이 3.80%이었음을 감안하면 본 연구에서 이용 한 분자마커 RM9와 벼 유전자원의 키다리병 저항성과는 불 일치성이 매우 높은 것을 확인할 수 있다. 감수성 품종 중 일 부 품종은 저항성 품종인 신광벼와 동일한 유전자형을 나타 내는 경우가 있으므로 신광벼의 저항성 QTL인
qBK1의연관 마커인 RM9는 다양한 유전자원 또는 교배후대에 대한 계통 선발 과정에서 범용적인 저항성 분자마커로 사용하기는 어려 울 것으로 사료되었다. 따라서 본 연구에서 선발한 저항성 유 전자원은 신광벼의 저항성 QTL인
qBK1와는 다른 영역 또는 염색체에 저항성 유전자가 존재할 가능성이 많다는 것을 시 사 하였다.
Fig. 3.Amplification pattern of the tightly linked marker RM9 among the resistant and susceptible varieties. Lane 1; Gwangmyeongbyeo, 2; Nep Huong, 3; Bukruksa209, 4; Misamgeum, 5; Nampyeongbyeo 6; Hawn, 7; Seonamna141, 8; Wonseadaesoo, 9; Erguailai, 10; Hanareum 2, 11; Namcheonbyeo, 12; Nogyang, 13; Dasanbyeo, 14; Junambyeo, 15; Boramchan, 16; Geurubyeo, 17; Hopum, 18; Seolgaeng
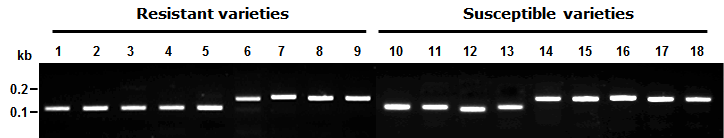
Table 3.Relationship between bakanae disease resistance and allele type of RM9, a bakanae disease resistance related marker, using 167 rice varieties
Table 3.
|
Phenotype |
Resistance |
Susceptible |
Mismatch ratio between allele type and phenotype (%) |
|
|
Allele type of RM9 |
Ilpum |
Shingwang |
Ilpum |
Shingwang |
|
|
Number of plants |
5 |
6 |
97 |
59 |
38.3 |
최근 벼 키다리병이 세계적으로 많이 발생되는 추세이나 저항성 관련 연구는 활발하지 않은 실정이다. 지금까지 벼 키 다리병에 대한 저항성 연구는 중국에서는 Chunjiang 06/TN1 DH 집단을 이용하여 1번 및 10번 염색체에서 저항성 QTL인
qB1과
qB10(
Yang et al. 2006)과 국내에서 일품/신광 BC
6F
3 집단을 이용하여 1번 염색체에 존재하는 신광 유래의 저항성 QTL인
qBK1(
Hur et al. 2015)이 밝혀진 바 있다. 다양한 저 항성 QTL의 탐색은 저항성 붕괴의 따른 대책의 일환인 gene pyramiding을 가능하게 하므로 저항성 유전자원을 이용한 추 가적인 저항성 QTL 연구가 필요하다. 본 연구에서는 병원성 균주인
G. fujikuroi CF283를 이용하여 생물검정을 수행하였 으나 벼 키다리병의 원인균인
Gibberella (
Fusarium) 속 곰팡 이는 그 종류가 다양하고 기주범위도 넓으므로 다양한 저항 성 유전자원의 확보와 저항성 유전자에 대한 연구를 위해서 는 추가적인 키다리병 균주에 대한 저항성 연구가 수행되어 야 할 것이다. 다양한 균주에 대한 생물검정 결과와 더불어 본 연구에서 선발된 저항성 유전자원과 기존에 선발된 저항 성 분자마커와의 상관성에 대한 정보를 통해 다양한 지역 및 환경조건 하에서 키다리병에 대한 저항성을 개선하는데 기여 할 수 있을 것으로 생각된다.
벼 키다리병은 Gibberella fujikuroi 또는 Fusarium moniliforme 등에 의해 발생하는 대표적인 종자전염성병으로, 육묘부터 수확 까지 지속적으로 발생하는 병해이다. 우리나라에서는 1960년 대 일부 농가에 심하게 발생하여 문제가 되었고, 최근 들어 벼 종자 내부의 심한 감염과 약제에 대한 저항성균의 출현 등 에 의하여 그 발병이 급격히 증가하여 보급종 종자에까지 발 병하고 있는 추세이다. 또한 벼 키다리병에 대하여 저항성이 높은 벼 품종이 개발되어있지 않아, 방제를 위한 농약살포에 막대한 농약과 노동력이 소모되고 있고, 이에 따라 생산비 절 감과 친환경 농산물의 생산에 큰 걸림돌이 되고 있는 실정이 다. 벼 키다리병 저항성 유전자는 1번 염색체에 존재하는 qB1 및 qBK1, 10번 염색체에 존재하는 qB10 등이 알려져 있다. 본 연구는 국내 육성 품종 및 국외 도입 유전자원을 중심으로 총 254 품종에 대해 생물검정을 통해 저항성 자원을 선발하 고, 기존에 개발된 신광벼의 저항성 QTL인 qBK1 영역의 분 자마커 RM9를 이용하여 유전적 다양성을 확인함을 목적으로 수행하였다. 본 연구에서는 중도 이상의 저항성을 보인 품종 은 11품종으로 전체 공시품종의 약 4%정도이었으며, 나머지 품종은 저항성 정도가 낮거나 약한 것으로 나타났다. 키다리 병 생물검정 결과와 분자마커 RM9에 대한 유전형과의 불일 치성은 38%로 나타났다. 이는 분자마커 RM9는 신광벼 후대 를 이용한 저항성 분자마커로 이용 가능하나, 다양한 유전자 원 또는 교배후대에 대한 범용적인 저항성 분자마커로 사용 하기에는 어려울 것으로 사료되었다. 특히, 본 연구에서 선발 한 저항성 유전자원은 향후 신광벼의 저항성 QTL인 qBK1외 의 새로운 는 다른 영역 또는 염색체에 저항성 유전자가 존재 할 가능성이 많다는 것을 시사 한다. 따라서 이들 품종은 유 전분석과 유전자지도작성을 통하여 키다리병 관련 저항성 신 규 저항성 유전자 여부에 대한 상세한 검토가 필요한 것으로 생각되며 향후 저항성 증진 또는 저항성 붕괴에 따른 대책으 로 gene pyramiding 등에도 이용 가능할 것으로 판단된다.
사 사
본 논문은 농촌진흥청 연구사업(세부과제명: 키다리병 저 항성 분자마커 개발을 통한 중간모본 개발, 세부과제번호: PJ011127022016)의 지원에 의해 이루어진 것임.
References
- 1. Carter LLA, Leslie JF, Webster RK. Population structure of Fusarium fujikuroi from California rice and water grass. Phytopathol 2008. 98: 992-998.
- 2. Chen DH, Ronard PC. A rapid DNA mini-preparation method suitable for AFLP and other PCR application. Plant Mol. Biol. Rep 1999. 17: 53-57.
- 3. Han S. Review of disease occurrence of major crops in Korea in 2007. Proceedings of Annual Falling Meeting and Symposium of KSPP 2007. pp 19-20.
- 4. Hori S. Researches on ‘bakanae’ disease of the rice plant. Nojishikenjyo Seiseki 1898. 12: 110-119.
- 5. Hur YJ, Lee SB, Kim TH, Kwon T, Lee JH, Shin DJ, Park SK, Hwang UH, Cho JH, Yoon YN, Yeo US, Song UC, Kwak DY, Nam MH, Park DS. Mapping of qBK1, a major QTL for bakanae disease resistance in rice. Mol. Breed 2015. 35: 78
- 6. Iqbal M, Javed N, Sahi ST, Cheema NM. Genetic management of bakanae disease of rice and evaluation of various fungicides against Fusarium moniliforme in vitro. Pakistan. J. Phytopathol 2011. 23: 103-107.
- 7. Khokhar LK, Jaffrey AH. Identification of sources of resistance against bakanae and foot rot disease in rice. Pakistan. J. Agric. Res 2002. 17: 176-177.
- 8. Kim MH, Hur YJ, Lee SB, Kwon TM, Hwang UH, Park SK, Yoon YN, Lee JH, Cho JH, Shin DJ, Kim TH, Han SI, Yeo US, Song YC, Nam MH, Park DS. Large-scale screening analysis for the evaluation of bakanae disease in rice. J. Gen. Plant. Pathol 2014. 80: 408-414.
- 9. Korea Crop Protection Association.Guideline for Pesticide Usage. 2011. pp. 288-292.
- 10. Kwon T, Lee JH, Park SK, Hwang UH, Cho JH, Kwak DY, Youn YN, Yeo US, Song YC, Nam JS, Kang HW, Nam MH, Park DS. Fine mapping and identification of candidate rice genes associated with qSTV11SG, a major QTL for rice stripe disease resistance. Theor. Appl. Genet 2012. 125: 1033-1046.
- 11. Lee YH, Kim SY, Choi HW, Lee MJ, Ra DS, Kim IS, Park JW, Lee SW. Fungicide resistance of Fusarium fujikuroi isolates isolated in Korea. Korean J. Pest Sci 2010. 14: 427-432.
- 12. Lee YH, Lee MJ, Choi HW, Kim ST, Park JW, Myung IS, Park K, Lee SW. Development of in vitro seedling screening method for selection of resistant rice against bakanae disease. Res. Plant Dis 2011. 17: 288-294.
- 13. Li DJ, Luo K, Chen Z. Studies on resistance of rice materials to bakanae disease and pathogenicity of pathogen (Fusarium moniliforme). Acta Phytopathol. Sinica 1993. 23: 315-319.
- 14. Lv B. Preliminary study of identification for resistance to rice bakanae disease in rice resource. Plant Protect 1994. 3: 20-21.
- 15. Ma L, Ji Z, Bao J, Zhu X, Li X, Zhuang J, Yang C, Xia Y. Response oof rice genotypes carrying different dwarf genes to Fusarium moniliforme and giberellic acid. Plant Prod. Sci 2008. 11: 134-138.
- 16. Mew TW, Gonzales PG. A handbook of rice seedborne fungi. 2002.
- 17. Ogawa K. Damage by “bakanae” disease and its chemical control. Jpn. Pestic. Inf 1988. 52: 13-15.
- 18. Ou SH. Rice diseases. IRRI, Los Banõs 1985.
- 19. Park WS, Choi HW, Han SS, Shin DB, Shim HK, Jung ES, Lee SW, Lim CK, Lee YH. Control of Bakanae Disease of Rice by Seed Soaking into the Mixed Solution of Procholraz and Fludioxnil. Res. Plant Dis 2009. 15: 94-100.
- 20. Rood MA. Bakanae in-field yield loss. Rice. J 2004. 15: 8-10.
- 21. Rosales AM, Mew TW. Suppression of Fusarium moniliforme in rice by rice-associated antagonistic bacteria. Plant Dis 1997. 81: 49-52.
- 22. Saremi H, Ammarellou A, Marefat A, Okhovvat SM. Binam a rice cultivar, resistant for root rot disease on rice caused by Fusarium moniliforme in Northwest, Iran. Int. J. Bot 2008. 4: 383-389.
- 23. Sukapanpotharan S, Chantarasnit A. A screening method for bakanae resistance. International Rice Research Newaletter 1980. 5: 6.
- 24. Sunder S, Virk KS. Studies on correlation between bakanae incidence and yield loss in paddy. Indian Phytopathol 1997. 50: 99-101.
- 25. Yamanaka S, Honkura R. Symptoms on rice seedlings inoculated with “Bakanae” disease fungus, Fusarium moniliforme Sheldon. Annals of the Phytopathol Society of Japan 1978. 44: 57-58.
- 26. Yang CD, Guo LB, Li XM, Ji ZJ, Ma LY, Qian Q. Analysis of QTLs for resistance to rice bakanae disease. Chin. J. Rice Sci 2006. 6: 657-659.
- 27. Yang H, Wang Z, Wu H, Zhu C. Study on the variation of rice bakanae under the different methods of seedling rising. J. Anhui Agri. Sci 2003. 31: 119-124.
- 28. Zainudin NIM, Izzati NA, Salleh B. Variability of Fusarium species associated with bakanae disease of rice based on their virulence, vegetative and biological compatibilities. Sydowia 2010. 62: 89-104.
Citations
Citations to this article as recorded by

- Genome-Wide Association Study to identify Bakanae disease resistance-related QTLs carrying novel candidate genes in rice (Oryza sativa L.)
Yuting Zeng, Fang-Yuan Cao, Ah-Rim Lee, Dongryung Lee, Backki Kim, Soon-Wook Kwon
npj Science of Plants.2025;[Epub] CrossRef - Map-Based Cloning and Characterization of a Major QTL Gene, FfR1, Which Confers Resistance to Rice Bakanae Disease
Hyeonso Ji, Kyeong-Seong Cheon, Yunji Shin, Chaewon Lee, Seungmin Son, Hyoja Oh, Dong-Kyung Yoon, Seoyeon Lee, Mihyun Cho, Soojin Jun, Gang-Seob Lee, Jeongho Baek, Song Lim Kim, Il-Pyung Ahn, Jae-Hyeon Oh, Hye-Jin Yoon, Young-Soon Cha, Kyung-Hwan Kim
International Journal of Molecular Sciences.2024; 25(11): 6214. CrossRef - Identification of qBK2.1, a novel QTL controlling rice resistance against Fusarium fujikuroi
Szu-Yu Chen, Ming-Hsin Lai, Yi-Ling Chu, Dong-Hong Wu, Chih-Wei Tung, Yue-Jie Chen, Chia-Lin Chung
Botanical Studies.2023;[Epub] CrossRef - Evaluation of Major Rice Varieties for Bakanae Disease Resistance in Korea
Sais-Beul Lee, Ju-Won Kang, Ji-Yoon Lee, Gi-Un Seong, Youngho Kwon, So-Myeong Lee, Nkulu Rolly Kabang, Jun-Hyeon Cho, Seong-Hwan Oh, Dongjin Shin, Jong-Hee Lee, Ki-Won Oh, Dong-Soo Park
Korean Journal of Breeding Science.2023; 55(2): 103. CrossRef - An explanation of the mystifying bakanae disease narrative for tomorrow's rice
Qaiser Shakeel, Mustansar Mubeen, Muhammad Aamir Sohail, Sajjad Ali, Yasir Iftikhar, Rabia Tahir Bajwa, Muhammad Anjum Aqueel, Sudhir K. Upadhyay, Praveen Kumar Divvela, Lei Zhou
Frontiers in Microbiology.2023;[Epub] CrossRef - Breeding of the High-Quality Medium-Late Maturing Rice Variety ‘Aram’ in Gyeongnam Province
Deok-Gyeong Seong, Young-Gwang Kim, Byeong-Han Lee, Jin-Woo Nam, Yeong-Min Jin, Yeon-Hyeon Hwang, Seong-Tea Lee, Su-Min Yun, Jeuong-Ju Lee, Jung-Sung Chung
Korean Journal of Breeding Science.2023; 55(4): 373. CrossRef - Past and Future Epidemiological Perspectives and Integrated Management of Rice Bakanae in Korea
Soobin Shin, Hyunjoo Ryu, Jin-Yong Jung, Yoon-Ju Yoon, Gudam Kwon, Nahyun Lee, Na Hee Kim, Rowoon Lee, Jiseon Oh, Minju Baek, Yoon Soo Choi, Jungho Lee, Kwang-Hyung Kim
The Plant Pathology Journal.2023; 39(1): 1. CrossRef - Current Studies on Bakanae Disease in Rice: Host Range, Molecular Identification, and Disease Management
Yu Na An, Chandrasekaran Murugesan, Hyowon Choi, Ki Deok Kim, Se-Chul Chun
Mycobiology.2023; 51(4): 195. CrossRef - A Novel Locus for Bakanae Disease Resistance, qBK4T, Identified in Rice
Sais-Beul Lee, Ji-Yoon Lee, Ju-Won Kang, Hyunggon Mang, Nkulu Rolly Kabange, Gi-Un Seong, Youngho Kwon, So-Myeong Lee, Dongjin Shin, Jong-Hee Lee, Jun-Hyeon Cho, Ki-Won Oh, Dong-Soo Park
Agronomy.2022; 12(10): 2567. CrossRef - Evaluation of the Rsistant to Bakanae Disease in Korean Rice Landraces (Oryza sativa L.)
Soon-Wook Kwon, Na-Eun Kim, Sang-Hyeon Jin, Jeonghwan Seo, Tae-Ho Ham, Joohyun Lee
Plant Breeding and Biotechnology.2021; 9(4): 355. CrossRef - Various Methods for Controlling the Bakanae Disease in Rice
Li Fang Jing, Haruhisa Suga
Reviews in Agricultural Science.2021; 9: 195. CrossRef - Mapping of a Major QTL, qBK1Z, for Bakanae Disease Resistance in Rice
Sais-Beul Lee, Namgyu Kim, Sumin Jo, Yeon-Jae Hur, Ji-Youn Lee, Jun-Hyeon Cho, Jong-Hee Lee, Ju-Won Kang, You-Chun Song, Maurene Bombay, Sung-Ryul Kim, Jungkwan Lee, Young-Su Seo, Jong-Min Ko, Dong-Soo Park
Plants.2021; 10(3): 434. CrossRef - Development of ‘MY299BK’, a Cultivar Resistant to Bakanae Disease Harboring qBK1 Gene Derived From a Tong-il Type Rice ‘Shingwang’
Sumin Jo, Sais-Beul Lee, Youn-Jae Hur, Ji-Yoon Lee, Jun-Hyeon Cho, No-Bong Park, Dong-Jin Shin, Jong-Hee Lee, Young-Bo Sohn, Snag-Ik Han, Seong-Hwan Oh, You-Chun Song, Dong-Soo Park
Korean Journal of Breeding Science.2020; 52(2): 172. CrossRef - Rice Genome Resequencing Reveals a Major Quantitative Trait Locus for Resistance to Bakanae Disease Caused by Fusarium fujikuroi
Do-Yu Kang, Kyeong-Seong Cheon, Jun Oh, Hyoja Oh, Song Lim Kim, Nyunhee Kim, Eungyeong Lee, Inchan Choi, Jeongho Baek, Kyung-Hwan Kim, Nam-Jin Chung, Hyeonso Ji
International Journal of Molecular Sciences.2019; 20(10): 2598. CrossRef - Kompetitive Allele-Specific PCR Marker Development and Quantitative Trait Locus Mapping for Bakanae Disease Resistance in Korean Japonica Rice Varieties
Kyeong-Seong Cheon, Young-Min Jeong, Youn-Young Lee, Jun Oh, Do-Yu Kang, Hyoja Oh, Song Lim Kim, Nyunhee Kim, Eungyeong Lee, Jeongho Baek, Inchan Choi, Kyung-Hwan Kim, Yong Jae Won, In Sun Yoon, Young-il Cho, Jung-Heon Han, Hyeonso Ji
Plant Breeding and Biotechnology.2019; 7(3): 208. CrossRef - Mapping of a major quantitative trait locus for bakanae disease resistance in rice by genome resequencing
Hyeonso Ji, Tae-Ho Kim, Gang-Seob Lee, Hyun-Ju Kang, Seung-Bum Lee, Seok Cheol Suh, Song Lim Kim, Inchan Choi, Jeongho Baek, Kyung-Hwan Kim
Molecular Genetics and Genomics.2018; 293(3): 579. CrossRef - Molecular mapping of qBK1WD, a major QTL for bakanae disease resistance in rice
Sais-Beul Lee, Yeon-Jae Hur, Jun-Hyeon Cho, Jong-Hee Lee, Tae-Heon Kim, Soo-Min Cho, You-Chun Song, Young-Su Seo, Jungkwan Lee, Tae-sung Kim, Yong-Jin Park, Myung-Kyu Oh, Dong-Soo Park
Rice.2018;[Epub] CrossRef