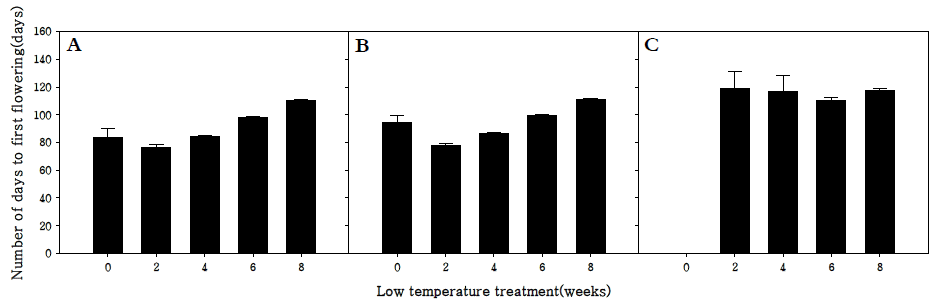적요
Flower bud differentiation by temperature and photoperiod was investigated in early-, medium-, and late-maturing cultivars of radish (Raphanus sativus L.). Flower bud differentiation was induced by only day-length extension treatment (18 hr light/day) in early - and medium-maturing cultivars. Therefore, the early- and medium-maturing cultivars of radish were advanced the time of the flowering by the 2 weeks low temperature duration with day-length extension (18 hr light/day). The vegetative growth of early- and medium-maturing cultivar was the best by only day-length extension (18 hr light/day). However, late-maturing cultivar was needed for the flower bud differentiation and optimal vegetative growth not only long-day but also low temperature during 4~6 weeks at least. Our results suggest the optimal condition for flower bud differentiation in the each early-, medium-, and late-maturing cultivars of radish, which will be useful to develop breeding materials of radish.
서 언
무(
Raphanus sativus L.)는 십자화과 채소로 우리나라의 김 치 주재료로 이용되고 있다. 무는 단백질, 지질, 당질, 섬유질, 비타민, 철, 마그네슘, 칼슘과 다양한 유용기능성물질인 디아 스타아제, 아밀라아제, 글루코시놀레이트를 함유하고 있어 식 용 및 약용으로 높은 영양학적 가치를 지니고 있으며, 우리나 라 7대 채소 중에 속할 만큼 중요한 위치를 차지하고 있으나 가격의 등락폭이 너무 심해 농가의 경제적 안정을 꾀할 수 없 는 상황이다(
Moon et al. 2012). 따라서 단순히 식재료로 이 용되던 무의 활용도를 높여 무 재배농가의 안정적인 소득증 대를 꾀하기 위하여 무에 함유된 기능성 물질을 극대화시킨 고품질의 신품종 육성이 절실히 필요한 상황이다. 또한 수확 대상물이 큰 무의 뿌리를 이용하여 유용기능성 물질 생산한 다면 원료 생산 및 수급이 원활할 것으로 기대된다. 이를 실 현하기 위해서는 유용기능성물질 함유량을 극대화시킨 고품 질 무의 육성해야 하는데 관행육종을 이용한다면 계통 육성 에 6 ~ 7년이 소요된다. 하지만 반수체 육종을 이용한다면 1 ~ 2년 사이에 계통 육성이 가능할 것으로 판단된다(
Jo et al. 2012,
Koh et al. 2010,
Kim et al. 2011). 이렇게 신품종 육 성을 위해 반수체 육종, 즉 소포자 및 약 배양이 시도되고 있 는데 아직 무의 경우 일부 제한된 품종 및 계통에서만 적용되 고 있는 상황이다(
Chun & Na 2011,
Chun et al. 2011). 하 지만 반수체 육종을 이용하여 신품종을 육성하기 위해서는 소포자 및 약 배양의 육종재료로 이용될 건전한 화뢰를 단기 간에 다량 획득하는 것이 중요하다. 무는 종자춘화형 식물로 저온에 감응되어야 화아분화가 이루어지는데, 화아분화를 위 한 저온감응도는 품종에 따라 다르다(
Moon et al. 2012). 그 러나 화아분화 유도에 요구되는 저온은 식물의 영양생장이 둔화된다는 문제점을 가지고 있다. 무의 화아분화에는 저온뿐 만 아니라 장일·강광 등의 환경요인이 작용한다는 보고가 있 으나(
Erwin et al. 2002) 각 품종에 따른 반응 및 효과는 구명 되지 않은 실정이다. 따라서 본 연구는 육종연한을 단축시키 고, 건전한 육종재료를 단기간에 획득하기 위해 무의 품종에 따른 화아분화 유도 기술을 확립하고, 저온 및 장일처리가 무 의 화아분화에 미치는 영향을 조사하고자 수행하였다.
재료 및 방법
공시재료로 조생품종인 만추무(대농씨앗, 대한민국), 중생 품종인 대금무(농원, 대한민국) 및 만생품종인 백옥무(농우바 이오, 대한민국)를 선정하였으며, 저온처리와 장일처리(18 hr light/day)에 따른 품종별 개화 반응 및 생육을 조사하였다. 2015년 4월부터 2015년 9월까지 목포대학교 부속농장 유리 온실 내에서 실험을 수행하였다. 4월 3일에 세 품종을 각각 50공 트레이에 2판씩 파종하여 발아하기 전까지 저면관수를 지속적으로 실시하였으며, 발아 후에는 1일 1회 두상 관수하 였다. 파종 후 5주 동안 유리온실 내 23.0/18.0±1.0°C(주/야 간) 온도조건에서 자연광을 이용하여 생육시킨 후 5월 8일에 cold lab chamber(JSSC-700C, JS RESEARCH, inc., 대한 민국)안에서 광량 5 ~ 10μmol·m-2·s-1, 광주기 18/6h(명/암), 온 도 4°C 조건으로 저온처리를 실시하였다.
세 품종을 0, 2, 4, 6, 8주 동안 각각 저온처리 한 후 10개 체씩 각각 화분(28.5 × 21.0 × 29.5cm3)에 정식하여 유리온실 에서 장일처리(18 hr light/day)를 하였다. 장일처리 조건은 낮에는 자연광을 이용하였으며 18시부터 24시까지 30~35μ mol·m-2·s-1 광량으로 장일처리 하였으며 기온은 23.0/18.0±1. 0°C(주/야간) 으로 설정하여 재배하였다. 토양은 시중에 유통 되고 있는 바이오상토(홍농종묘(주))를 사용하였다.
생육조사는 첫 개화일로부터 일주일 지난 식물체를 수확하 여 초장, 엽면적, 엽수, 지상부와 지하부의 생체중 및 건물중 을 측정하였고, 각 식물체마다 첫 개화시기를 조사하였다. 엽 면적과 엽수는 엽장이 1.0 cm 이상인 잎을 채취하여 생육조 사를 실시하였으며, 엽면적은 엽면적계(Li-3100C, LI-COR, Inc., USA)로 측정하였다. 조사 식물체의 건물중은 자연순환 식 건조기(HB-501M, Hanbaek Co. Ltd., 대한민국)를 이용 하여 75°C에서 5일 동안 건조한 후 중량을 측정하였다.
결과 및 고찰
저온처리 및 장일처리(18 hr light/day)에 따른 화아분화 양상을 조사한 결과 조생종과 중생종은 장일처리(18 hr light/day)만으로도 화아분화가 가능한 반면 만생종은 장일처 리(18 hr light/day)로 화아분화를 유도할 수 없었다(Fig.
1). 조생종과 중생종의 경우 2주 동안 저온처리를 하였을 때 첫 개화까지 소요되는 기간이 각각 76.5, 78.2일로 가장 짧게 소 요되었으며 저온처리를 하지 않고 장일처리(18 hr light/day) 만으로 화아분화를 유도한 처리구는 각각 83.4, 94.4일로 개 화까지 소요되는 기간은 2주 동안 저온처리 한 처리구보다 5 ~ 6일 정도 지연되었으나 화아분화는 정상적으로 유도되는 것을 확인하였다. 하지만 만생종의 경우 6주 동안 저온처리를 한 처리구가 첫 개화까지 110일이 소요되는 반면 160일 이상 장일조건에서 생육시켜도 화아가 전혀 분화되지 않는 것을 확인하였다(Fig.
1). 이는 무의 춘화처리는 장일보다는 저온 이 더 효과적이지만 장일조건에 장시간 노출될 경우 화아분 화를 유도할 수 있다는 기존의 이론에 동의는 되지만 만생종 의 경우에는 이 이론에 해당되지 않음을 확인하였다(
Yoo & Andrew 1984). 조생종과 중생종은 저온처리기간이 길어질수 록 개화시기가 늦어지는 결과를 나타내었다. 백합과에 속하는 비비추의 경우 저온처리를 실시할 경우 저온처리시기, 저온처 리기간과는 무관하게 맹아와 개화를 유도할 수 있었으나 저 온처리를 대신하여 장일처리(18 hr light/day)를 할 경우에는 기간과 시기에 민감한 반응을 보였는데 이는 저온처리에서는 품종에 상관없이 개화하지만 일장처리의 경우에는 품종 및 처리기간에 민감하게 반응하는 무와 비슷한 경향을 가진 것 으로 사료된다(
Jeong 2005). 기존에는 무의 화아분화를 유도 하기 위해 4 ~ 6주 동안 저온처리를 실시하였는데, 본 연구 결 과를 보면, 조생종과 중생종의 경우 저온처리를 대신하여 장 일처리(18 hr light/day)만으로 개화를 유도할 수 있을 뿐 아 니라 2주 동안의 저온처리를 실시함으로써 개화소요기간을 최소화시킬 수 있을 것으로 판단된다. 본 연구 결과는 각각의 품종에 적합한 저온처리기간을 구명하여 시간과 노동력, 그리 고 에너지를 절약하는데 효과적으로 이용될 수 있을 것으로 사료된다.
Fig. 1.Effect of periods low temperature treatment on the number of days to flowering in R. sativus. Early-maturing cultivar (A), medium-maturing cultivar (B), late-maturing cultivar (C).
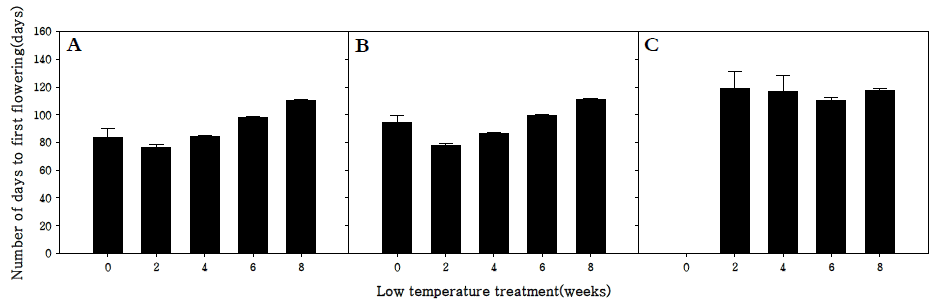
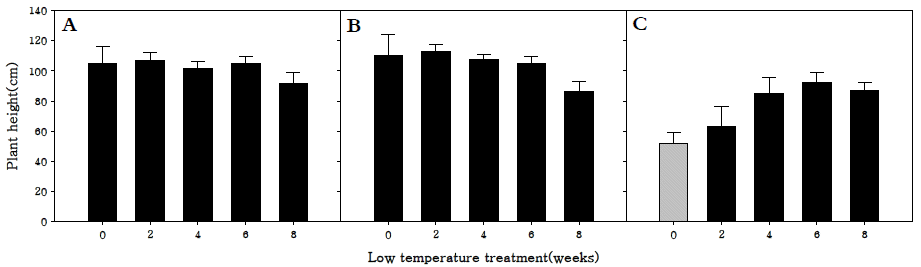
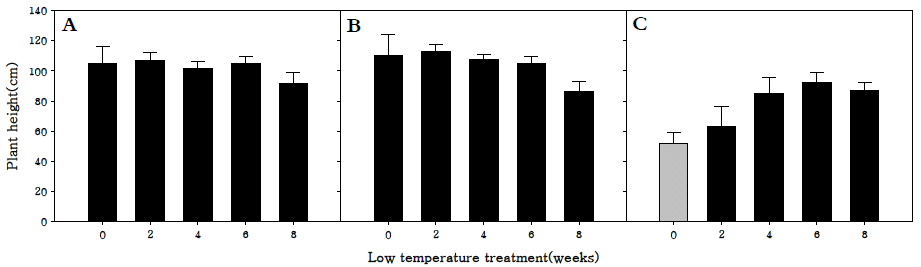
화아분화 유도 후의 초장을 조사한 결과, 조생종과 중생종 의 경우 2주 동안 저온처리를 하였을 때 개화 소요기간을 가 장 단축시켰을 뿐 아니라 유의한 차이를 보이지 않았지만 107.1, 113.2 cm로 초장이 가장 긴 것을 확인하였다. 또한 8 주 동안 저온처리를 한 처리구는 다른 처리구에 비해 유의하 게 초장이 짧았다(Fig.
2). 이는 충분이 저온에 감응한 후 고 온 장일에 노출되어야 개화와 추대가 진행되는데, 저온 감응 후에도 저온에 노출되어 있어 생육이 지연되었을 것으로 판 단된다(
Moon & Yu 2013). 만생종의 경우 6주까지는 저온처 리 기간이 증가할수록 초장도 증가하는 경향을 보였다. 만생 종은 조생종과 중생종에 비교하여 개화에 요구되는 저온요구 도가 더 큰 것으로 나타났으며, 따라서 저온에 충분히 감응한 후 고온장일에 노출된 식물체가 추대를 함으로써 초장이 증 가하는 것으로 판단된다.
Fig. 2.Length of aboveground parts in R. sativus. Early-maturing cultivar (A), medium-maturing cultivar (B), late-maturing cultivar (C).
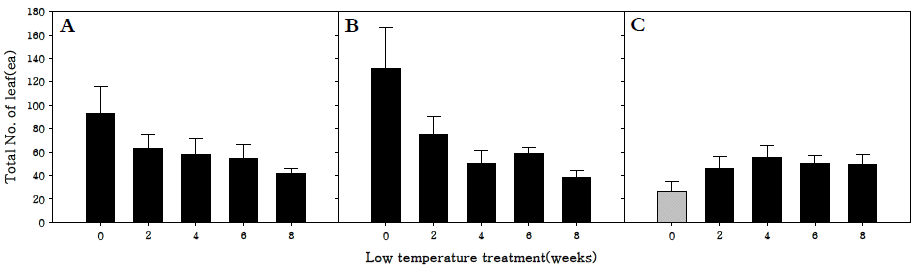
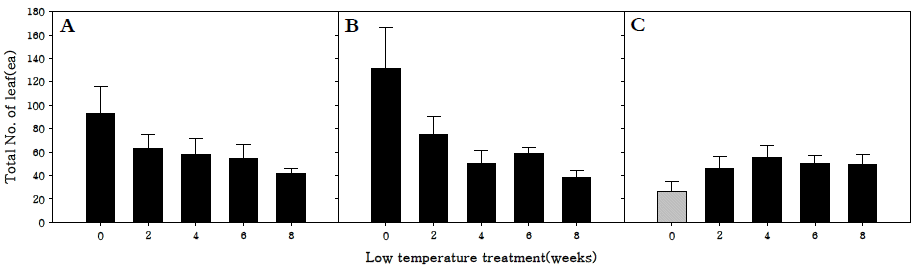
저온처리 기간에 따른 엽수를 조사한 결과, 조생종과 중생 종은 저온 0주 처리구에서 각각 93.3, 131.6개로 가장 많았다 (Fig.
3). 이는 저온처리 대신 장일·강광 조건을 부여함으로써 저온으로 인한 생육둔감현상이 감소하였으며, 화아분화 이전 에 최적 생육 조건에 노출됨으로 영양생장이 촉진된 것으로 판단된다.
Fig. 3.Total number of leaf in R. sativus. Early-maturing cultivar (A), medium-maturing cultivar (B), late-maturing cultivar (C).
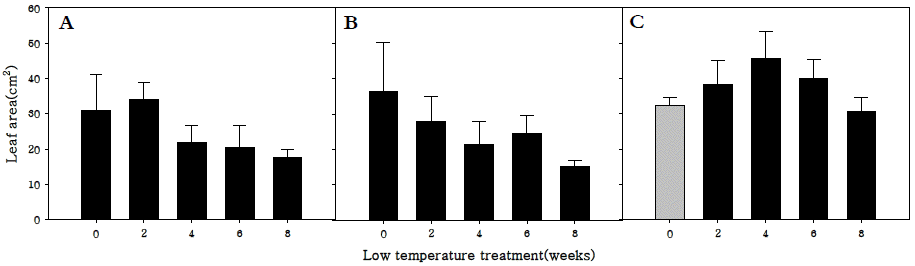
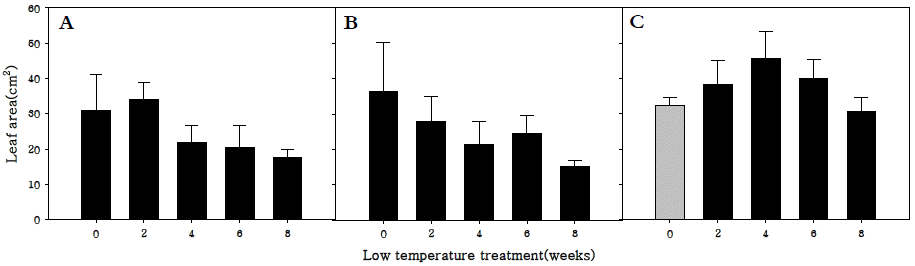
엽면적을 조사한 결과, 조생종과 중생종의 경우 저온처리 기간이 증가할수록 엽면적은 감소하는 경향을 나타내었으나 만생종의 경우 0주에서부터 4주 동안 저온처리할 경우는 저 온저장 기간이 증가할 수록 엽면적이 증가하였다가 4주 이상 부터는 저온처리기간이 증가할수록 감소하는 경향을 보였다 (Fig.
4). 이는 저온요구도가 가장 높은 만생종의 경우 저온처 리가 무의 화아분화 뿐 아니라 생육에도 밀접하게 관계가 있 음을 나타내는 결과로 사료된다.
Fig. 4.Leaf area in R. sativus. Early-maturing cultivar (A), medium-maturing cultivar (B), late-maturing cultivar (C).
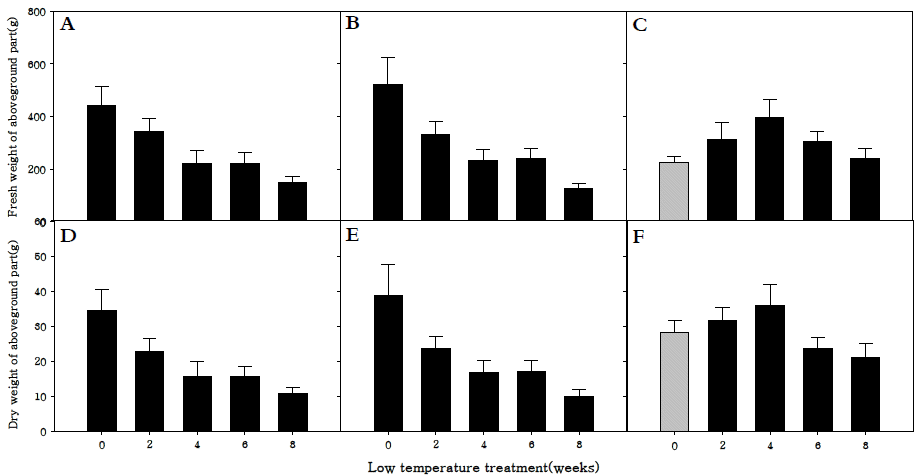
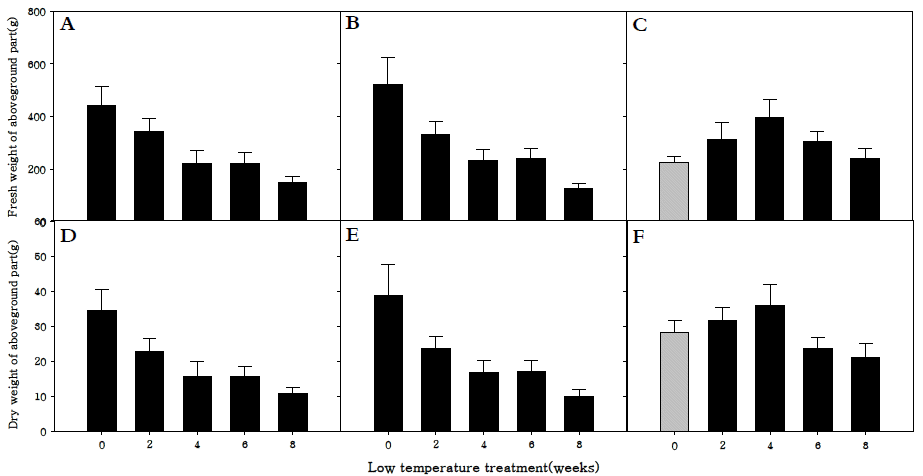
지상부 생체중과 건물중을 조사한 결과, 조생종과 중생종 의 생체중과 건물중은 저온 0주 처리구에서 가장 높았다(Fig.
5A-
F). 또한 저온처리 기간이 길어질수록 생체중 및 건물중 이 감소하는 경향을 보였다. 조생종과 만생종은 저온처리를 하지 않고 강광·장일조건에서 생육시킴으로써 영양생장기관 을 충실하게 생육시킬 수 있었으며, 충실한 영양생장 후 생식 생장으로 전환시켜 충실하고 건강한 화뢰를 얻을 수 있었던 것으로 판단된다. 만생종은 엽면적 결과처럼 저온 4주 처리구 에서 생체중과 건물중 모두 가장 증가하였다. 이 결과로 인하 여 만생종의 경우 저온에 충분히 감응시키는 것이 화아분화 뿐 아니라 지상부의 생육에도 효과적이라는 것을 확인할 수 있었다.
Fig. 5.Fresh weight of aboveground parts (A, B, C) and dry weight of aboveground parts (D, E, F) in R. sativus. Early-maturing cultivar (A, D), medium-maturing cultivar (B, E), late-maturing cultivar (C,F).
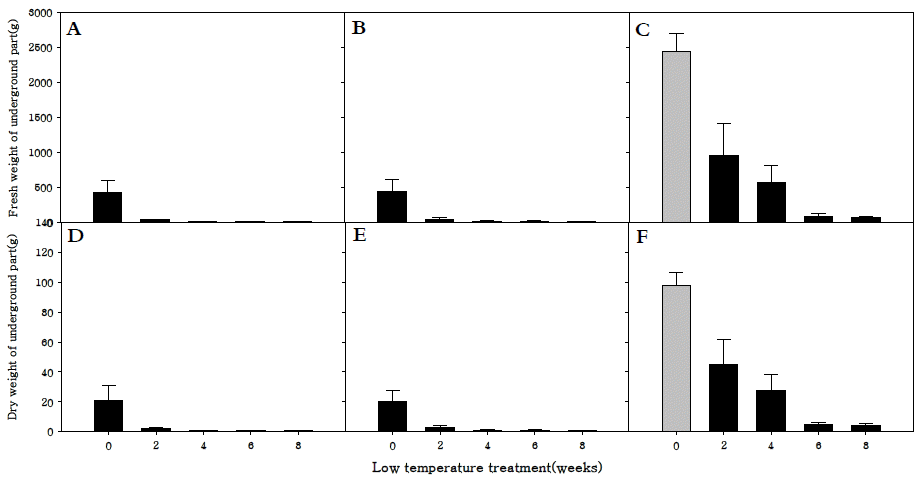
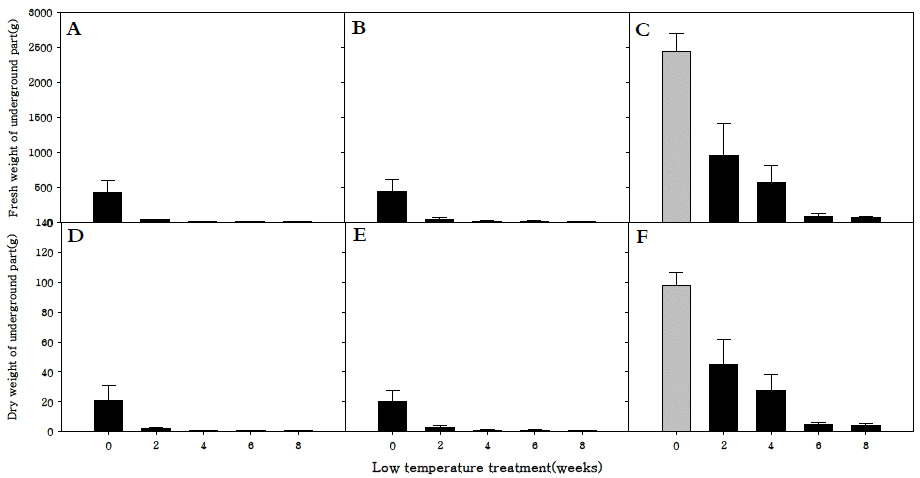
지하부 생체중과 건물중은 세 품종 모두 저온처리를 하지 않았을 때 가장 높았으며, 특히 만생종은 조생종과 중생중에 비해 월등히 높은 것을 확인하였다(Fig.
6A-
F). 이는 화아분 화가 진행되지 않고 영양생장기관만 생장하던 만생종은 화아 분화가 진행되어 양분의 흡인이 생식생장으로 이동된 조생종, 중생종보다 지하부의 생육이 더 왕성하게 진행되었기 때문으 로 사료된다.
Fig. 6.Fresh weight of underground parts (A, B, C) and dry weight of underground parts (D, E, F) in R. sativus. Early-maturing cultivar (A, D), medium-maturing cultivar (B, E), late-maturing cultivar (C,F).
본 연구를 통해 무의 조생종과 중생종은 저온처리를 생략 하고 강광·장일처리(18 hr light/day)만으로 화아분화 유도 가 가능하지만 만생종의 경우는 저온처리가 꼭 필요한 것을 확인하였으며, 또한 개화소요기간을 최소로 단축하기 위한 각 품종에 따른 최적 저온처리기간을 구명하였다.
적 요
무의 조생종, 중생종, 만생종에 저온 및 장일 처리가 화아 분화 및 생육에 미치는 영향을 구명하기 위해 실험을 수행한 결과, 조생종과 중생종은 장일처리(18 hr light/day)를 하는 것만으로도 화아분화 유도가 가능하였으며 2주 동안 저온처 리를 하였을 때 개화소요기간이 가장 짧았다. 하지만 만생종 의 경우 장일처리(18 hr light/day)만으로는 화아분화 유도가 불가능하였으며 6주 동안 저온처리를 하였을 때 개화까지 소 요되는 기간이 가장 짧았다. 또한 조생종과 중생종의 경우 장 일처리(18 hr light/day)만을 하였을 때 모든 영양기관의 생육 이 가장 좋았으며, 만생종의 경우 지하부 생육을 제외한 모든 영양기관의 생육은 저온처리를 4 ~ 6주 동안 하였을 때 가장 효과적이었다. 조생종과 중생종의 경우 저온처리를 하지 않고 장일처리(18 hr light/day)만으로 건전한 식물체로의 생육뿐 아니라 화아분화 유도가 가능하였으나, 만생종의 화아분화에 는 저온처리가 필수요인임을 확인하였다. 이 연구를 통하여 무의 각 품종에 적합한 화아분화 유도 조건을 구명하였으며, 본 연구결과는 단기간에 육종소재를 개발하는데 이용 가능할 것으로 기대된다.
사 사
본 논문은 2013학년도 목포대학교 교내연구비 지원에 의 하여 연구되었음
References
- 1. Chun CH, Na HY. Microspore-derived embryo formation in response to cold pretreatment, washing medium, and medium composition of Radish (Raphanus sativus L.). Kor. J. Hort. Sci. Technol 2011. 29: 494-499.
- 2. Chun CH, Park HY, Na HY. Microspore-derived embryo formation in Radish (Raphanus sativus L.) according to nutritional and environmental conditions. Hort. Environ. Biotechnol 2011. 52: 530-535.
- 3. Erwin JE, Warner RM, Smith AG. Vernalization, photoperiod and GA3 interact to affect flowering of Japanese radish (Raphanus sativus Chinese Radish Jumbo Scarlet). Physiol. Plant 2002. 115: 298-302.
- 4. Jeong JH. Effects of low temperature and photoperiod treatment on growth and flowering in Hosta longipes. Kor. Ass. Flo. Ind. Dev 2005. 13: 273-278.
- 5. Jo MH, Ham IK, Park MY, Kim TI, Lim YP, Lee EM. Seed production ability of doubled haploid plants through microspore culture in Chiness Cabbage (Brassica rapa L. ssp. pekinensis) introduced from China. Kor. J. Hort. Sci. Technol 2012. 30: 573-578.
- 6. Kim YJ, Lee KW, Kim HS, Cheong YK, Park KG, Kim KJ. Effects of plant growth regulators on haploid production of wheat. Korean J. Breed. Sci 2011. 43: 50-55.
- 7. Koh GH, Kim EH, Park EH, Seo YW, An SL. Plant breeding science. 2010. Hayng munsa, Seoul, Korea: pp. 407-410.
- 8. Moon W, Kim JK, Lee JW. Horticultural crop science I - vegetable. 2012. Knou, Seoul, Korea: pp. 123-357.
- 9. Moon W, Yu DJ. Crop physiology science. 2013. Knou, Seoul, Korea: pp. 325-334.
- 10. Yoo KC, Andrew WT. Studies on the physiology of bolting and flowering in Raphanus sativus L. VIII. Effect of irradiant energy and day length on the vernalized and non vernalized Radishes. Kor. J. Hort. Sci. Technol 1984. 25: 201-205.
Citations
Citations to this article as recorded by

- The Molecular Mechanism of Relatively Low-Temperature-Induced Broccoli Flower Bud Differentiation Revealed by Transcriptomic Profiling
Wenchen Chai, Xia He, Boyue Wen, Yajie Jiang, Zixuan Zhang, Rui Bai, Xinling Zhang, Jin Xu, Leiping Hou, Meilan Li, Jing Zhang
Horticulturae.2023; 9(12): 1353. CrossRef - Appropriateness Evaluation of Plug Seedling Cultivation for Replanting of Rehmannia glutinosa (Gaertn.) Libosch. ex Steud
Sang Hoon Lee, Seung Uk Kang, So Hee Lee, Sung Cheol Koo, Mok Hur, Mei Lan Jin, Woo Moon Lee, Min Su Park, Yeon Bok Kim, Jong Won Han
Korean Journal of Medicinal Crop Science.2019; 27(3): 202. CrossRef