적요
Antigen production in plant is a safe and effective strategy for vaccine development. In this study, rice transformants were developed for oral vaccine against pigs diarrhea disease. DNA cassette composed with the cholera toxin subunit B (CTB) connected to the 987P-fasG, for stimulating a strong oral immune response, was introduced to rice through Agrobacterium mediated genetic transformation. Copy number analysis by TaqMan real-time PCR for transgenes revealed that transgene of 1 to 8 copies have been introduced into T1 and T2 rice seeds. The expression level of mRNA in the transformants T1 and T2 generations were up to 35 times higher than the reference value in the result of analysis by Quantitative real time-PCR. In addition, the callus cultured from rice transformants was confirmed that the introduced gene has been maintained till 9-month subculture duration. The amount of mRNA expression value was also confirmed in callus, which was maintained above 2.6 times compared with that of the standard control for a long time. These results provide that the introduced antigen for plant-based vaccine against bacterial diarrhea disease can be maintained in the callus as well as in the transgenic plant and suggest that the callus culture of plant transformant will be an effective way to obtain a plant-derived edible vaccine.
서 언
경구용 식물 백신 개발은 병원체의 항원유전자를 식물에 도입 시킴으로써 식물체가 항원단백질을 생산하게 하는 기술인데, 식물섭식을 통해 치료가 이루어지므로 기존의 백신에 비해 거부 감이 적으며 식물생산체계를 이용한 대량생산이 가능하므로 생산비용이 저렴할 뿐만 아니라 보관이 용이하다는 장점 등이 있다(
Youm et al. 2010). 그러나 발현량의 정도, 형질전환체를 사용함으로 인한 환경위해성 등의 문제점도 있어 해결되어야 할 과제 또한 많다고 하겠다. 현재 단백질 발현 수준을 높이기 위해 여러 가지 프로모터에 관한 연구가 진행 중이며 최근에는 식물 바이러스 벡터 시스템과 아그로박테리움을 이용한 agroinfiltration을 같이 사용하는 시스템이 외래단백질 발현을 높인다는 연구결과가 보고되기도 하였다(
Hefferon 2014).
현재 개발이 진행중인 식물유래 재조합 항원은 CTB(cholera toxin B subunit), LTB(heat labile enterotoxin B subunit), HBV(hepatitisB surface antigen), HPV(Human papillomavirus), HIV(Human immunodeficiency virus) 등이 있으며, 사용되는 식물도 감자, 담배, 토마토, 벼, 콩, 상추, 바나나, 당근 등으로 다양하다(
Thanavala et al. 2010,
Nozoye et al. 2009,
Moravec et al. 2007,
Kohl et al. 2007,
Ramirez et al. 2007). 특히 돼지 설사병 등에 관여하는 CTB는 그 동안 감자, 토마토 등에 형질전환으로 도입된 바 있으며,
Nochi et al.(2007)은 형질전환 벼 유래의 CTB는 실온에서 장기간 보관되어도 활성이 유지되므 로 백신으로의 활용이 가능하다고 하였다. 쌀, 옥수수, 콩 등 곡물을 사용하면 단백질을 종자에 축적할 수 있어 안전하게 저장할 수 있을 뿐만 아니라 넓은 면적에서 대량생산이 가능하므 로 가축의 사료로 이용시 경제적이라 하겠다(
Lamphear et al. 2002).
본 연구에서는 돼지 설사병을 예방하는 경구용 식물 백신 개발의 일환으로 콜레라 독소 항원 유전자를 생산하는 벼 형질전 환체를 개발하였으며, 또한 형질전환체 유래 캘러스를 장기간 보관하며 도입유전자의 발현을 분석하였기에 이를 보고하고자 한다.
재료 및 방법
형질전환체 개발
돼지설사병 방제용 식물경구백신을 개발하고자 관련 유전자 를 이용하여 벼를 형질전환시켰으며, 특히 식물경구백신이 보다 효율적으로 면역작용을 촉진시킬 수 있도록 점막면역보조제인 Cholera toxin subunit B (CTB) 유전자와 돼지의 설사병을 유발시키는 장독소 대장균 987P-fasG 유전자를 연결시킨 융합 백신을 개발하고자 하였다. 점막면역보조제와 항원 유전자가 융합된
CTB::fasG 유전자는 단국대학교(
Hwang, 2008)에서 분 양받아 사용하였다. 클로닝된
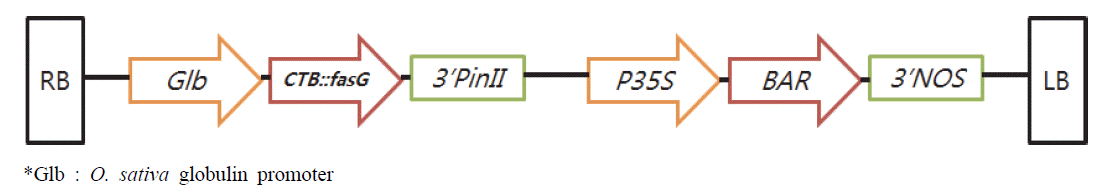
CTB::fasG DNA 카세트는 단자엽 식물 형질전환에 사용되며 벼 글로불린 프로모터(Glb)를 지니고 있는 pMJ103 vector에 재조합(Fig.
1) 시킨 후, 형질전환을 위해
Agrobacterium tumefaciens LBA4404에 도입시켰다. 식물 형질전환의 재료는 일미벼 종자에서 유래된 캘러스를 사용하였 다.
Agrobacteria와 캘러스를 4일간 28°C 암 조건으로 공동배양 한 후 L-phosphinotricine(PPT, 6mg/L)과 cefotaxime(250mg/ L)이 포함된 N6 배지에 치상하고 27°C 암 조건에서 배양하며 형질전환된 캘러스를 선발하였다. 선발된 캘러스는 PPT 3mg/L, naphthalene acetic acid(NAA) 1mg/L, kinetin 5mg/L이 포함 된 MS 배지에 치상한 다음 25°C의 명조건에서 배양하며 shoot 를 분화시켰으며, 재분화 식물체는 생장호르몬이 포함되지 않은 MS배지에 옮겨 뿌리를 유도하고 형질전환체 T
0를 양성하였다. 이후 T
0 개체는 자가수분 시켜 T
1 세대를 양성하였으며, PCR을 통하여 유전자가 도입된 것이 확인된 T
1 세대를 자가수분 시켜 T
2 세대로 후대를 진전시키고 실험재료로 사용하였다.
Fig. 1.Map of the T-DNA region of recombinant vector pMJ 103 harboring CTB::fasG.
CTB::fasG 유전자 도입 확인
형질전환체의
CTB::fasG 유전자 도입 확인은 T
1 및 T
2 종자를 대상으로 PCR(Polymerase Chain Reaction)로 수행하였다. 각 종자는 액체질소를 이용하여 막자 사발로 분쇄하고 CTAB법으 로 gDNA를 추출한 후 template로 사용하였다. Primer는
CTB::fasG가 증폭되도록 디자인 한 Not I CF forward와 reverse primer를 10pmol의 농도로 조정하여 사용하였다(Table
1). PCR은 LaboPass
™ SP-Taq PCR Master mix(Cosmo Genetech., Korea)를 사용하였으며, 94°C 5분; 94°C 20초, 60°C 40초, 72°C 100초, 30cycles; 72°C 5분의 조건으로 수행하였다. 한편, T
1 및 T
2종자에서 조직배양을 통하여 얻어진 callus를 대상으로도 유전자 도입을 확인하였다. Callus는 2,4-D가 2mg 첨가된 2N6 배지, 25°C, 암배양 조건에서 배양하였다. 유도된 캘러스는 1 개월마다 동일한 배지에 계대배양 하였으며 1, 2, 3, 6, 7, 8, 9개월간 배양된 캘러스는 수확 후 액체질소를 이용하여 분쇄한 후 CTAB법으로 gDNA를 추출하고 종자의 경우와 같은 방법으로 PCR을 수행하였다.
Table 1.List of primers used in this study.
Table 1.
|
Primer |
Sequence (5' - 3') |
|
|
NotI CF F |
TTGCGGCCGCAAATGATCAAGCTCAAGTTTGGAGT |
|
NotI CF R |
TTGCGGCCGGAACTAACCTTTTTCCCATCTGCTTA |
|
|
|
Nos F |
GCATGACGTTATTTATGAGATGGGTTT |
|
Nos R |
TGCGCGCTATATTTTGTTTTCTATCG |
|
Forward probe |
TAGAGTCCCGCAATTAT |
|
|
|
CTB::fas G F |
CACCCAAATCCATACCC |
|
CTB::fas G R |
CCTTCATCCTCTCAATGG |
|
|
|
OsActin F |
ACAGGTATTCTCTTGGACTCTGG |
|
OsActin R |
AGTAACCACGCTCCGTCAGG |
형질전환 종자의 도입유전자 copy number assay
형질전환 T
1및 T
2종자를 대상으로 도입유전자 copy수를 확 인하기 위해 TaqMan probe법으로 Real-time PCR을 수행하였 다. 공시재료는 T
1 종자 5 lines, T
2 종자 8 lines이며 negative control로는 일미벼 종자를, positive control로는 2 copies로 계산된
nos gene을 사용하였다. CTAB법으로 추출한 gDNA 30ng과 TaqMan® Gene Expression Master Mix (Applied Biosystems, U.S.A)를 95°C, 10분; 94°C, 15초; 60°C, 60초 40cycles 조건으로 ABI StepOnePlus
™ (Applied Biosystems, U.S.A)을 사용하여 Real-time PCR을 수행하였다. Endogenous control은 rice
tubulin alpha-1 chain gene (AK102560)으로
nos gene의 forward, reverse primer와 forward probe를 사용하 였다(Table
1). 모든 실험은 3반복으로 수행되었으며, 외래 유전 자의 copy number 계산은 Applied Biosystems CopyCaller® Sofrware v2.0 (Applied Biosystems, U.S.A)을 이용하였다.
형질전환체에 도입된
CTB::fasG 유전자의 발현 수준을 분석 하기 위하여 Quantitative real time-PCR(qRT-PCR)를 수행하 였다. 공시재료는 온실에서 약 3개월 생장한 T
1및 T
2개체의 지상부와 조직배양 후 수확된 캘러스(1, 2, 3, 6, 7, 8, 9개월 배양)가 사용되었다. Total RNA는 Trizol®Reagent (Invitrogen, USA)를 사용하여 추출하였고, DiaStar
™ 2X RT Primix (SolGent, Korea)를 이용하여 total RNA 1
μg으로부터 first strand cDNA를 합성하였다. 합성된 cDNA는 SYBR Premix Ex Taq (Takara, Japan)과 함께 95°C, 30초; 95°C, 5초, 60°C, 34초 40cycles 조건으로 ABI StepOnePlus
™ (Applied Biosystems, U.S.A)을 사용하여 qRT-PCR 분석을 하였으며, 모든 실험은 3반복으로 수행되었다. Primer는
CTB gene의 일부가 증폭되게 디자인 한 CTB::fasG forward, reverse primer와 OsActin forward, reverse primer를 20 pmol이 되게 조정하여 사용하였다. Threshold cycle (Ct)값에 의한 발현량 분석은
Livak & Schmittgen (2001)의 2
-ΔΔCt 방법으로 계산하 였다.
결과 및 고찰
형질전환체의 CTB::fasG 유전자 도입 확인
형질전환체 T
1및 T
2개체는 생육시 제초제인 0.7% 바스터 처리를 통하여 삽입유전자가 도입되었음을 간접적으로 확인할 수 있었다. 또한 자가수분 후 수확된 종자에서
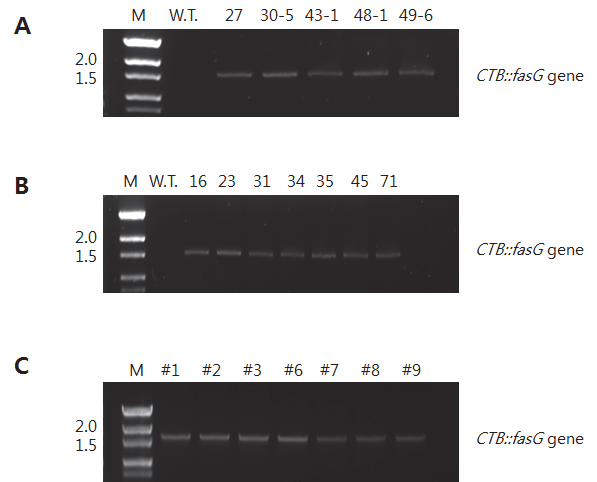
CTB::fasG 유전자 도입을 확인하기 위해 PCR을 수행하였다. 예상되는 PCR product 크기는 1,576bp이었으며 1% agarose gel에 전기영동 한 결과, 예상했던 크기에서 band가 확인되었다(Fig.
2-A and
B). Wild type인 일미벼 종자에서는 band가 나타나지 않았으나 공시된 T
1line 5개와 T
2line 7개 모두에서
CTB::fasG 유전자 크기의 밴드를 확인할 수 있었다.
Fig. 2.
PCR analysis of CTB::fasG gene in rice transformants. (A) T1progeny, (B) T2progeny, and (C) Callus of T149-6 line. Numeric of lane shows subcultured months.
*M: 100bp DNA ladder; WT: wild type.
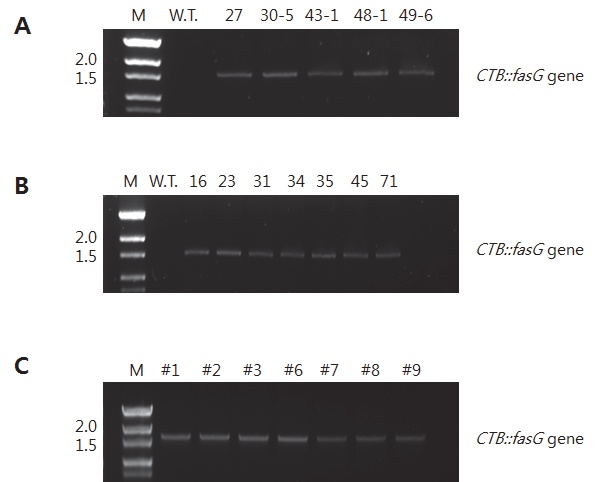
형질전환체를 조직배양한 캘러스에서 도입유전자가 발현되 는 지를 확인하였다(Fig.
2C). T
149-6 line의 종자를 조직배양하 여 유도된 캘러스에서 gDNA를 추출하여 PCR을 수행하였다. 캘러스는 1, 2, 3, 6, 7, 8, 9, 및 11 개월간 동일 배지에서 계대배양 하며 생육을 유지시켰다. Fig.
2C에서 보는 바와 같이 9개월간 기내배양에서 유도된 캘러스 모두에서 약 1.5kb의 band가 확인 되었다. 따라서 생육기간이 연장된 캘러스에서도
CTB::fasG 유전자가 유지된다고 판단할 수 있었다. 그런데 11 개월간 계대 배양한 캘러스에서는 PCR 결과 실험이 수행된 3 반복 모두에서 유전자가 확인되지 않았다. 이러한 결과가 장기간 배양에 따른 캘러스의 퇴화 때문인 지의 여부는 추후 좀 더 많은 연구가 있어야 할 것으로 사료된다. 그러나 캘러스에서의 도입유전자 유지 는 계대배양 기간 등이 어느 정도 영향을 끼칠 것으로 판단된다.
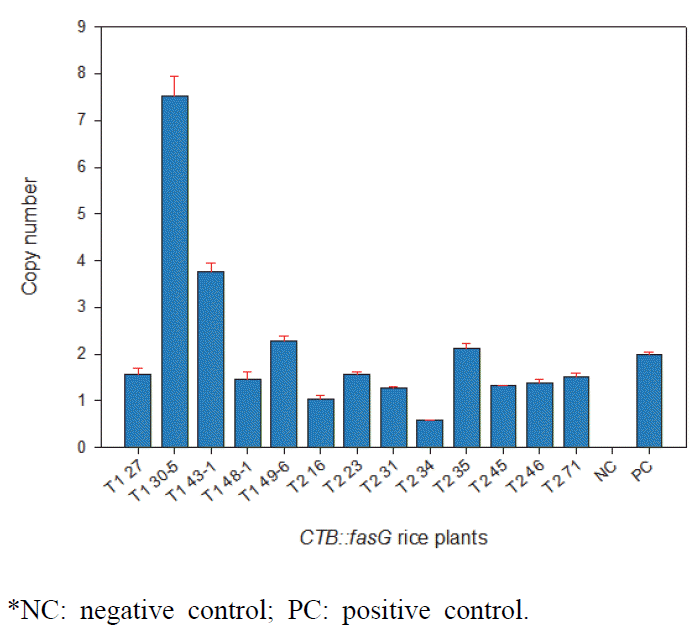
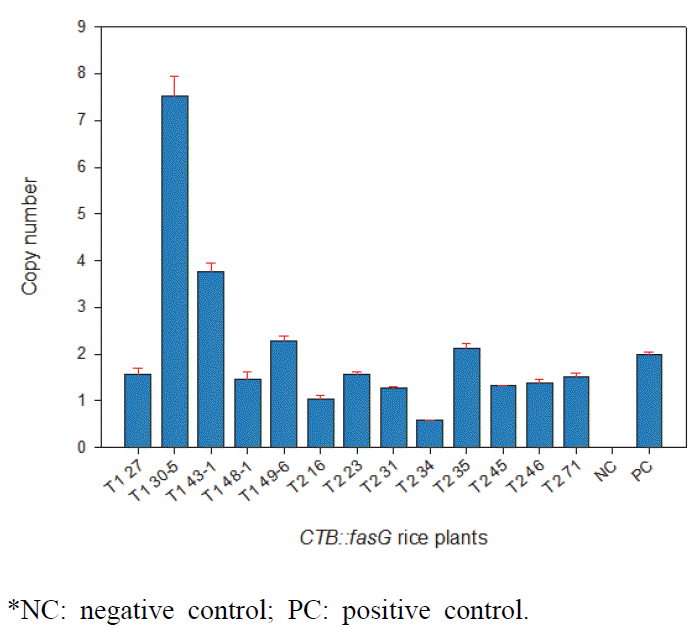
한편, 형질전환체에 도입된 유전자의 copy 수를 확인하기 위해 gDNA를 추출하여 TaqMan probe법으로 real-time PCR 을 수행하였다(Fig.
3). Positive control은 2 copies로 계산된
nos gene으로 하였으며 T
1 종자 5 lines, T
2 종자 8 lines을 대상으로 도입유전자 copy 수를 분석하였다. Negative control 인 일미벼 종자에서는 유전자 도입을 확인할 수 없었으나 형질전 환 종자들에서는 도입유전자의 copy 수가 1 ~ 8 개의 변이를 보이며 나타났다.
Fig. 3.Copy number assay in transgenic rice by quantitative RT-PCR.
형질전환 벼의 CTB::fasG mRNA 발현 분석
형질전환 벼의 지상부를 대상으로
CTB::fasG 유전자의 발현 수준 분석을 Quantitative real time-PCR(qRT-PCR)로 수행하 였다. 발현양은
Livak & Schmittgen (2001)의 2
-ΔΔCt법에 따라 threshold cycle(Ct)값에 의해 분석되었다. ΔCt는
CTB::fasG 유전자의 Ct값에서 reference gene으로 사용된
OsActin 유전자 의 Ct값을 뺀 것이며 ΔΔCt는
CTB::fasG의 ΔCt값 중 제일 높은 것을 reference로 하여 각 시료들의 ΔCt값에서 reference 의 ΔCt값을 뺀 값으로 분석되었다.
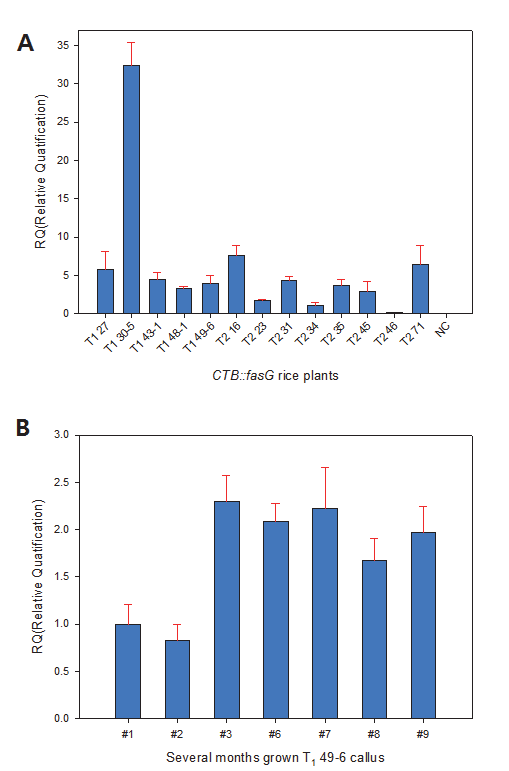
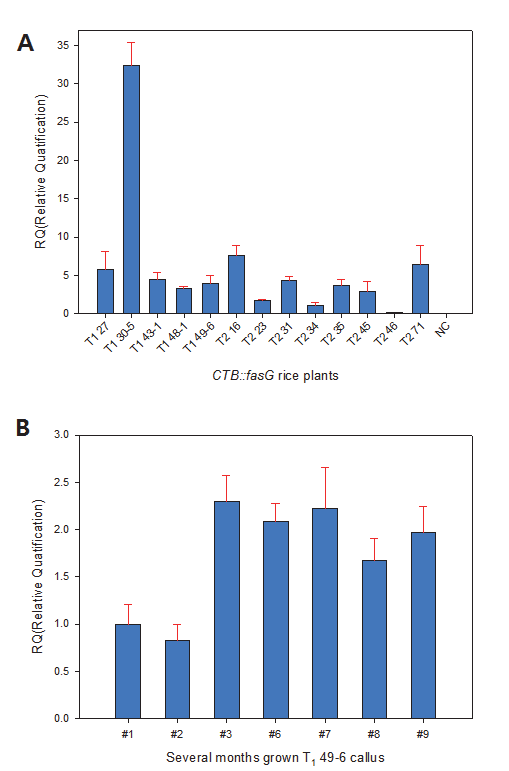
Fig.
4A는 형질전환 T
234 line의 분석치를 reference로 지정 하여 2
-ΔΔCt값을 구한 다음 다른 개체들과의 상대적인 mRNA 발현량을 비교한 결과이다. 그림에서 보는 바와 같이 negative control인 일미벼는
CTB::fasG 유전자의 mRNA가 발현되지 않음이 확인되었다. 형질전환 개체간 비교에서는 T
246은 RQ값 이 0에 가까워 T
2-34와 비교할 때 mRNA 발현이 거의 없었던 반면, T
130-5는 약 35배 가까이 발현되는 것으로 나타났다. T
271, T
216, T
127 등도 T
234에 비해 약 8배가 발현되었다. 한편, 도입유전자 copy 수와 mRNA 발현량간의 일정한 경향은 나타나지 않았다. 이것은 도입유전자 수는 종자에서 분석된 반면 mRNA 발현량은 지상부를 대상으로 분석되었기에 조직 및 기관 에 따라 mRNA가 다르게 발현되었을 것으로 생각된다. 그러나 도입유전자 수가 가장 크게 나타난 T
130-5는 mRNA 발현량도 다른 lines 에 비해 특이하게 높게 나타났으므로 도입유전자 수와 mRNA 발현량간에는 어느 정도 상관이 있으리라 판단된다.
Fig. 4.Expression analysis of the CTB::fasG mRNA by quantitative real time-PCR. OsActin was used as internal control. (A) Transgenic rice (T1 and T2progeny). (B) Callus of T149-6 line. Numeric of lane shows subcultured months.
따라서 본 연구에서 개발된 형질전환체를 이용하여 품종이 육성되고 이를 사료 등으로 사용할 수 있다면 어느 정도 돼지 설사병 예방에 도움이 될 수 있으리라 사료된다. 물론 형질전환 벼에서의 발현량이 실제 임상치료에 가능할 정도로 충족될 수 있을지 여부는 추후 연구가 더 필요할 것이다. 또한 본 실험에서 는 mRNA 발현량만을 분석하였기에 mRNA 발현량이 실질적인 단백질 발현과 상등한 결과를 보일 지에 대해서는 판단하기 곤란하다. 따라서 추후 단백질 발현 및 효능 분석에 관한 심도있 는 연구가 계속되어야 할 것이다.
형질전환 종자 T
149-6 line으로 수개월 조직배양한 캘러스를 대상으로
CTB::fasG mRNA 발현량을 분석하였다(Fig.
4B). 2개월 배양된 캘러스의 분석치를 기준으로 정하여 상대적인 발현량을 비교분석한 결과 7개월 배양한 캘러스의
CTB::fasG mRNA가 2.66배로 상대적으로 제일 많은 것으로 나타났다. 1개월 배양한 캘러스는 1.2배로 나타났으며 9개월 배양 캘러스 에서는 기준에 비해 2배 이상 mRNA가 발현되었다. 이와 같이 동일 line의 캘러스에서 발현량에 변이가 나타나는 것은 계대배 양 배지가 고체인 까닭에 캘러스의 축적상태가 상이한 것 등이 변이의 요인이 되었을 것으로 사료된다. 그러나 장기간 배양된 캘러스에서 mRNA가 지속적으로 발현된다는 것은 확실하게 판단할 수 있었다.
Hwang et al.(2015)은 황금(
Scutellaria baicalensis) 캘러스를 재료로 황금 함유 활성물질인 baicalein 등의 함유량을 분석한 결과 캘러스가 1년 정도 오랜 기간 배양되어도 함유 성분이 유지되며 배지 종류에 따라 함유량의 변이가 나타난다고 보고한 바 있어 본 연구결과를 뒷받침한다고 하겠다. 그런데 외래유전자 가 도입된 식물형질전환체의 캘러스를 재료로 도입유전자의 유지를 확인한 연구결과는 아직 없으므로 본 연구 결과는 상당히 주목할 사항이라 사료된다. 특히 캘러스 배양은 조절된 환경하에 서 대량번식이 가능할 뿐만 아니라 생장조절제 등 여러 요인의 변화로 필요성분 축적량을 높일 수 있는 방안이 될 수 있으므로 추후 관련연구가 지속되어야 할 것이다. 또한 현재 대두되고 있는 GM 작물 재배의 환경위해성 문제를 극복할 수 있는 하나의 방편이 될 수도 있을 것이라 판단된다.
적 요
돼지설사병 방제용 식물경구백신을 개발하고자 점막면역보 조제인 Cholera toxin subunit B (CTB) 유전자와 설사병을 유발시키는 대장균 987P-fasG 유전자를 연결시킨 CTB::fasG DNA 카세트로 벼를 형질전환시켰다. 벼에 삽입된 도입유전자 의 copy수를 T1및 T2세대의 종자를 대상으로 TaqMan probe법 으로 분석한 결과 1 개부터 8 개까지의 유전자가 복수로 도입된 것으로 확인하였다. 형질전환체 T1및 T2세대에서의 mRNA의 발현량은 Quantitative real time-PCR로 분석한 결과 기준치에 비해 최대 35배까지 나타났다. 또한 형질전환체를 조직배양하여 생성된 callus를 장시간 계대배양시키며 저장기간 별로 도입유전 자를 확인한 결과 계대배양 9개월까지 도입유전자가 유지됨을 확인할 수 있었다. Callus에서의 mRNA의 발현량 역시 기준치의 2.6배 정도로 유지됨을 확인하였다. 이와 같이 도입유전자를 형질전환체 뿐만 아니라 callus에서도 확인할 수 있었으므로 본 연구 결과는 식물유래 경구백신 개발의 기초자료로 활용될 수 있을 것으로 판단된다.
사 사
본 논문은 순천향대학교의 연구 지원에 의하여 수행되었습니다.
References
- 1. Arntzen C, Plotkin S, Dodet B. Plant-derivedvaccines and antibodies. Vaccines 2005. 23: 1753-1756.
- 2. Daniell H, Singh ND, Mason H, Streatfield SS. Plant-made vaccine and biopharmaceuticals. Trends Plant Sci 2009. 14(12):669-679.
- 3. Hwang IT, Lee JJ, Lee JY, Baek SW, Kim YH. Production of baicalin Baicalein and wogonin by plant tissue culture of Scutellaria baicalensis. Kor. J. Plant Resources 2015. 28(4):526-432.
- 4. Hwang CH. Development of plant oral vaccine against piglet bacterial diarrhea. RDA Report-20050401034714 2008.
- 5. Hefferon KL. DNA virus vectors for vaccine production in plants: spotlight on geminiviruses. Vaccines 2014. 34: 642-653.
- 6. Kim TG, Yang MS. Current status in molecular farming. J. Plant Biotechnology 2010. 37(3):243-249.
- 7. Kohl TO, Hitzeroth II, Christensen ND, Rybicki EP. Expression of HPV-II L1 protein in transgenic Arabodopsis thaliana and Nicotiana tabacum. BMC Biotechnol 2007. 12: 7-56.
- 8. Kwon JY, Cheon SH, Lee HR, Han JY, Kim DI. Production of biopharmaceuticals in transgenic plant cell suspension cultures. J. Plant Biotechnology 2009. 36(4):309-319.
- 9. Lamphear BJ, Streatfield SJ, Jilka JM, Brooks CA, Barker DK, Tumer DD, Delaney DE, Garcia M, Wiggins B, Woodard SL, Hood EE, Tizard IR, Lawhom B, Howard JA. Delivery of subunit vaccines in maize seed. J. Control Release 2002. 13: 169-180.
- 10. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔC` method. Methods 2001. 25: 402-408.
- 11. Moravec T, Schmidt MA, Herman EM, Woodford TT. Production of Escherichila coli heat labilet toxin(LB) B subunit in soybean seed and analysis of its immunogenicity as an oral vaccine. Vaccine 2007. 19: 1647-1657.
- 12. Nochi T, Takagi H, Yuki Y, Yang L, Masumura T, Mejima M. Rice-based mucosal vaccine as a global strategy for cold-chain-and needle-free vaccination. PNAS USA 2007. 104(26):10986-109911.
- 13. Nozoye T, Takaiwa F, Tsuji N, Yamakawa T, Arakawa T, Hayashi Y, Matsumoto Y. Production of Ascaris Suum As14 protein and its fusion protein with cholera toxin B subunit in rice seeds. J. Vet. Med. Sci 2009. 71: 995-1000.
- 14. Ramirez YJ, Tasciotti E, Gutierrez-Ortega A, Donayre Torres AJ, Olivera Flores MT, Giacca M, Gomez Lim MA. Fruit-specific expression of the human immunodeficiency virus type 1 tat gene I tomato plants and its immunogenic potential in mice. Clin. Vaccine Immunol 2007. 14: 685-692.
- 15. Sala F, Rigano MM, Barbante A, Basso B, Walmsley AM, Castiglione S. Vaccine antigen production in transgenic plants : strategies gene constructs and perspectives. Vaccines 2003. 21: 803-808.
- 16. Streatfield SJ. Plant-based vaccines for animal health. Rev. sci. tech. Off. int. Epiz 2005. 24(1):189-199.
- 17. Thanavala Y, Lugade AA. Oral transgenic plant-based vaccine for hepatitis B. Immunol. Res 2010. 46: 4-11.
- 18. Youm JW, Jeon JH, Joung H, Kim HS. Plants as platforms for the production of vaccine antigens. J. Plant Biotechnology 2010. 37(3):250-261.