적요
Chlorophyll is an essential pigment involved in light absorption and electron transfer in photosynthesis, a photochemical process that is indispensable for plant growth and development. The biosynthesis of chlorophyll occurs in plastids and shares a common biosynthetic pathway with other tetrapyrroles. The chlorophyll metabolic pathway is divided into four distinct components: the common pathway, chlorophyll-specific biosynthetic pathway, chlorophyll cycle, and chlorophyll degradation pathway, which are regulated in developmental- and environmental-specific manners. During the early stages of plant growth, the expression of most chlorophyll biosynthetic genes is induced by light, resulting in an increase in chlorophyll accumulation, induction of high photosynthetic activity, and continuous plant growth. In contrast, during plant maturation, the expression of most of these genes is gradually downregulated, whereas genes involved in chlorophyll degradation are upregulated during leaf senescence. Chlorophyll biosynthesis is directly or indirectly regulated by the members of various transcription factor families. In this review, we describe representative mechanisms of transcription factor-mediated activation and repression of chlorophyll biosynthesis in response to light treatment. We also present an overview of recent studies that have examined all the enzymatic steps involved in chlorophyll metabolic pathways and their gene regulation at the transcriptional level, which will enable readers to gain a better understanding of chlorophyll metabolism.
서 언
엽록소는 빛 에너지를 흡수하는 식물 색소로 광합성의 최종 산물인 탄소화합물 생성에 중요한 역할을 수행한다. 엽록소는 빛에 의해 유도되고, 유식물체에서 생성이 촉진되어 성숙시기까지 축적되며, 식물이 노화됨에 따라 분해가 점차적으로 증가되어진다. 이러한 엽록소는 테트라피롤(tetrapyrrole) 화합물 중 하나이며, 테트라피롤은 4개의 탄소와 1개의 질소로 이루어진 피롤 구조 4개가 일렬 또는 고리모양으로 연결된 화합물이다. 테트라피롤 화합물에는 엽록소 뿐 만 아니라, 헴(heme), 시로헴(siroheme), 및 피토크로모빌린(phytochromobilin) 등이 있다.
엽록소 생합성경로는
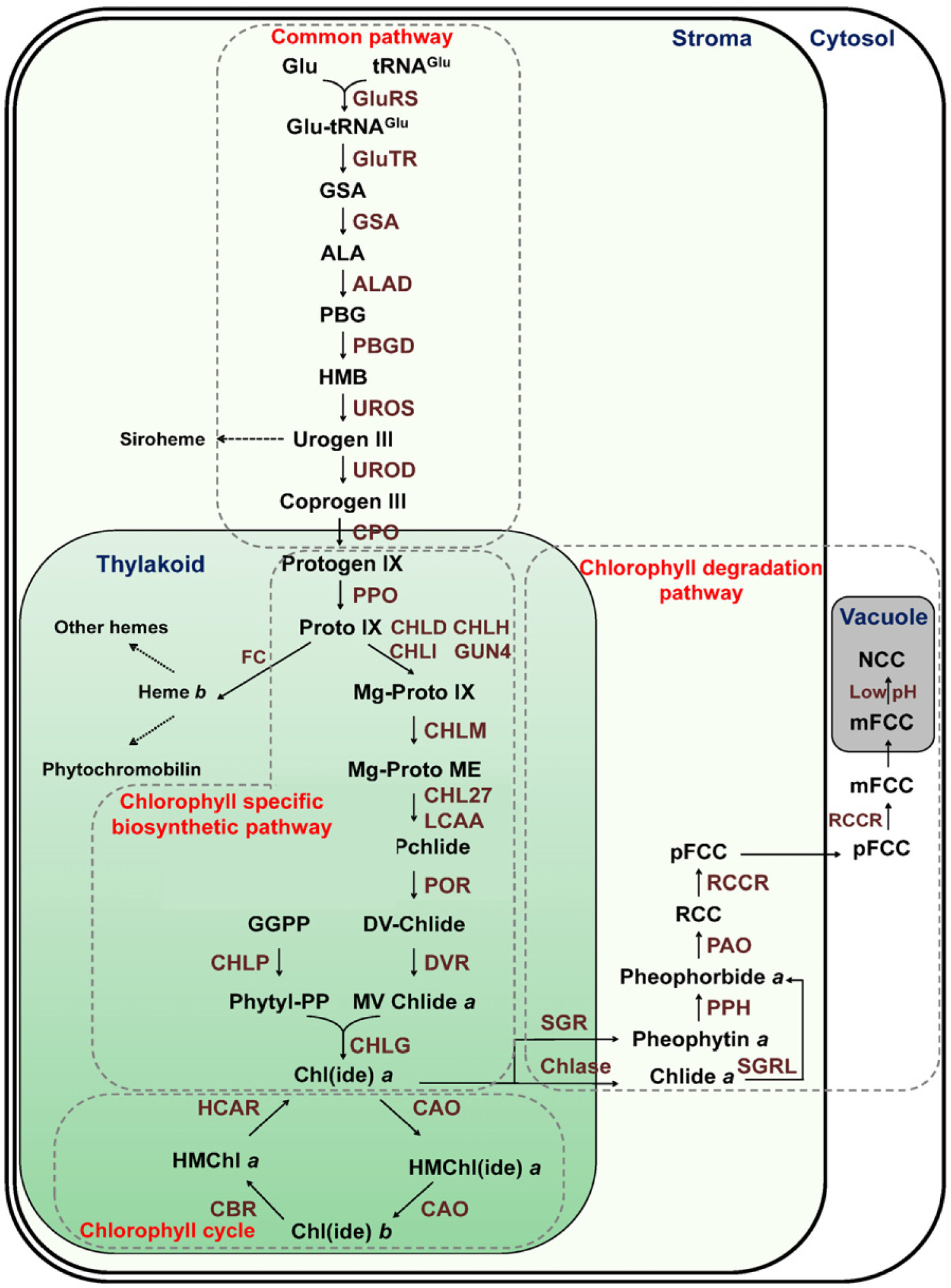
Fig. 1에서처럼 4단계로 나누어서 살펴볼 수 있다. 엽록소 대사는 테트라피롤계 물질이 생성되는 공통 경로, 엽록소가 생성되는 엽록소 특이적 생합성 경로, 엽록소
a (chlorophyll
a, Chl
a)와 엽록소
b (chlorophyll
b, Chl
b)가 상호 전환되는 엽록소 순환 경로, 그리고 최종적으로 엽록소가 분해되어 액포에 저장되는 분해 경로로 나뉜다. 엽록소는 식물의 광합성에 필수적인 물질이지만, 엽록소 생합성 경로에서 생성되는 대부분의 엽록소 중간물질들은 빛 조건하에서 일중항산소(singlet oxygen)를 쉽게 생성한다. 일중항산소는 식물의 잎에서 상시적으로 생산되며 프로그램된 세포 사멸을 유도하는 활성 산소의 일종이다. 식물은 엽록소 중간 물질들에 의한 일중항 산소의 생산을 억제하기 위하여 생장 동안 엽록소 생합성 경로를 엄격하게 규제한다(
Triantaphylidès & Havaux 2009). 또한, 엽록소 생합성에 관여하는 효소 활성은 보조인자, 산화환원반응, 피드백 시스템 등을 통하여 미세하게 조절되어진다(
Brzezowski et al. 2015,
Kobayashi & Masuda 2016). 본 논문에서는 엽록소 생합성 경로의 유전자들에 대한 기능을 설명하고, 이를 조절하는 전사 인자들을 소개하며 관련한 최근 연구들을 기술하고자 한다.
엽록소 생합성 경로
엽록소 대사에 관여하는 효소는 엽록체내에서 서로 다른 공간에 위치하여 효소 반응을 조절한다. 엽록소 대사의 초기단계인 공통 경로에 관여하는 효소들은 스트로마에 위치하여 엽록소 생성 뿐만 아니라 헴을 포함한 다양한 테트라피롤계 물질의 생성을 촉매하며, 엽록소 생합성 및 순환 경로를 촉매하는 효소들은 틸라코이드에 존재한다. 엽록소 분해 경로에 관여하는 효소들은 대사의 경로에 따라 각각 틸라코이드, 스트로마, 세포질에 위치하여 엽록소의 분해를 유도한다. 또한, 엽록소 생합성 경로에 관여하는 다수의 유전자는 일주기, 생물학적 주기에 의해서 조절되어진다. 엽록소 생합성 경로의 전반적인 과정은
Fig. 1에 도식화 하였으며, 쌍자엽과 단자엽의 대표적 모델 식물인 애기장대와 벼에서의 엽록소 생합성 경로에 관여하는 유전자들은
Table 1에 정리하였다.
엽록소 생합성의 첫 단계로 테트라피롤계 물질의 전구체인 아미노레불린산(5-aminolevulinic acid, ALA)의 생성에 있어서 세 가지 효소 단계가 요구된다(
Oborník & Green 2005). Glutamyl-tRNA synthetase (GluRS)에 의해 글루타메이트(glutamate, Glu)와 tRNA
Glu로부터 형성된 Glu-tRNA
Glu는 Glu-tRNA reductase (GluTR)에 의해 Glu 1-semialdehyde (GSA)로 환원된다. GluTR을 암호화하는 3개의 유전자(
HEMA1,
HEMA2,
HEMA3)가 애기장대에서 보고되었고(
McCormac et al. 2001), 이들은 서로 다른 발현 양상을 보인다. 애기장대의
HEMA1은 빛에 의해 유도되어 모든 조직에서 발현되며,
HEMA2는 뿌리에서 특이적으로 발현되고,
HEMA3는 기능이 없는 위유전자(pseudogene)로 보고되었다(
McCormac et al. 2001).
GluTR에 의해 생성된 GSA는 GSA aminotransferase (GSA)에 의해 ALA로 전환되어지며, 애기장대에서는 2개의
GSA 유전자가 이 과정에 관여하는 것으로 보고되었다(
Ilag et al. 1994). 두 분자의 ALA는 ALA dehydratase (ALAD)의 효소작용으로 비대칭적으로 응축되어 monopyrrole porphobilinogen (PBG)를 형성한다. PBG 분자 4개가 PBG deaminase (PBGD)에 의해 순차적으로 중합되어 불안정한 선형 테트라피롤인 hydroxymethylbilane (HMB)가 생성된다. 애기장대에는 1개의
PBGD 유전자를 지니고 있으며, 잎과 뿌리에서 발현한다(
Lim et al. 1994). HMB는 uroporphyrinogen III synthase (UROS)에 의해 유로포르피리노겐 III (uroporphyrinogen III, Urogen III)로 전환되며, 생성된 Urogen III의 일부는 다른 테트라피롤계 물질인 시로헴의 생성에도 이용된다(
Tanaka et al. 2011). 애기장대 유래의
UORS를 대장균에 도입한 뒤 단백질을 분리하여 효소 활성을 분석한 결과, 정상적인 UROS는 전구체인 HMB를 Urogen III로 전환시켰으나, 염기서열이 치환된 UROS 돌연변이의 경우 HMB가 비효소적으로 고리화 되어 네 번째 피롤 고리의 재배열 없이 Urogen I을 형성하였다. Urogen I은 생물학적 테트라피롤의 전구체가 아니며, 경로의 다음 단계로 나아갈 수 없다(
Tan et al. 2008).
Urogen III는 Urogen III decarboxylase (UROD)에 의해 코프로포르피리노겐 III (coproporphyrinogen III, Coprogen III)로 변환된다. Coprogen III는 Coprogen III oxidase (CPO)에 의해 산화되어 프로토포르피리노겐 IX (protoporphyrinogen IX, Protogen IX)를 형성한다. 애기장대에서는 두 개의
CPO 유전자가 보고되었으며, 돌연변이체 분석을 통하여
CPO1은 기능을 하는 유전자이며(
Ishikawa et al. 2001),
CPO2는 위유전자로 보고되었다(
Matsumoto et al. 2004).
Protogen IX는 Protogen IX oxidase (PPO)에 의해 추가로 산화되어 엽록소와 헴 생성의 전구체인 프로토포르피린 IX (protoporphyrin, Proto IX)를 생성한다. 애기장대에는 2개의
PPO 유전자가 존재하며,
PPO1은 빛에 의해 발현이 유도되며 기능을 하는 유전자이고,
PPO2는 발현이 거의 되지 않는다(
Zhang et al. 2014). Proto IX의 형성 후, 공통 경로는 중심 이온이 마그네슘(Mg
2+)인 엽록소 생합성 경로와 중심 이온이 철(Fe
2+)인 헴 생합성 경로로 나누어 진다.
Proto IX는 Mg-chelatase에 의해 ATP 의존적 방식으로 Mg
2+이온이 삽입되어 Mg-Proto IX를 생성한다. Mg-chelatase는 Mg-chelatase subunit D (CHLD), Mg-chelatase subunit H (CHLH), Mg-chelatase subunit I (CHLI) 서브유닛으로 구성되어 있으며, CHLD와 CHLI는 복합체를 형성하여 Mg
2+를 공급하는 반면 CHLH는 기질을 수송하는 것으로 보고되었다(
Kobayashi et al. 2008). Genomes uncoupled 4 (GUN4)는 막 내에서 Mg-chelatase 복합체중 CHLH와 결합하여 엽록소 생합성을 활성화시킨다. 다른 엽록소 생합성 경로의 유전자들과 다르게
GUN4는 엽록소 축적에 필수적이지는 않지만, 중간 물질의 흐름을 조절함으로써 Mg-chelatase의 활성을 돕는다(
Adhikari et al. 2011,
Davison et al. 2005). 암조건시 GUN4의 C-말단이 인산화 되어짐에 따라 효소 활성이 불활성화되어 Mg-chelatase의 활성을 촉진하지 못하여, 엽록소의 생성은 줄어들고 헴 생성이 늘어나게 된다. 반대로 빛 조건시 GUN4에 의해 Mg-chelatase가 활성화되고, 엽록소 생성이 증가되어진다. 이처럼 빛에 의한 GUN4의 활성화에 따라서 식물체 내의 헴 생성과 엽록소 생성 간의 균형이 유지된다(
Richter et al. 2016).
Mg-chelatase에 의해 생성된 Mg-Proto IX는 Mg-Proto IX methyltransferase (CHLM)에 의해 메틸기가 추가되어 Mg-Proto IX monomethyl ester (Mg-proto ME)로 전환되며, Mg-Proto ME cyclase에 의해 3,8-divinyl-protochlorophyllide (DV-Pchlide)로 전환된다. Mg-Proto ME cyclase는 막 결합 서브 유닛인 CHL27 (
Rzeznicka et al. 2005)과 low Chl accumulation A (LCAA) 서브 유닛의 복합체로 구성된다(
Albus et al. 2012,
Hollingshead et al. 2012). 분자생물학적 분석을 통해 CHL27과 LCAA의 직접적인 상호작용이 확인되었다(
Albus et al. 2012). 덱사메타손-유도(dexamethasone-induced) RNAi (RNA interference) 실험을 통한
CHL27의 일시적인 발현 감소는 Mg-Proto IX와 Mg-Proto ME간의 양적 불균형을 발생시켜 빛 조건시 유해 활성 산소를 생성시켰다(
Schlicke et al. 2014). 또한, RNAi에 의한
LCAA 발현 억제 담배 식물체에서도 Mg-Proto ME의 축적이 확인되었으며, DV-Pchlide 함량 감소와
CHL27 발현 감소가 확인되었다(
Schlicke et al. 2014).
NADPH: Pchlide oxidoreductase (POR)에 의해 DV-Pchlide의 D고리가 환원되어 3,8-divinyl-chlorophyllide (DV-Chlide)로 전환된다. 빛 의존적 POR은 혐기성 박테리아를 제외한 모든 광합성 유기체에서 발견되었다. 반면, 빛 독립적 POR의 경우에는 PORA, PORB, 및 PORC 세 개의 서브유닛으로 구성되어 있으며 모든 겉씨식물, 호기성 광합성 박테리아 및 녹조류에 존재하지만, 속씨식물에는 존재하지 않는다. 속씨식물의 경우에는 암조건에서 빛 독립적인 POR이 존재하지 않음으로 엽록소 생성이 일어나지 않으며, DV-Pchlide를 축적한다(
Eckhardt et al. 2004). 애기장대에는 3개의
POR 유전자가 존재하며,
PORA는 암조건에서 발현이 유도되는 반면,
PORB는 모든 조직에서 상시적으로 발현이 되고,
PORC는 빛에 의해 발현이 유도된다(
Eckhardt et al. 2004).
DV-Chlide는 DV-Chlide reductase (DVR)에 의해 3-vinyl Chlide
a (MV-Chlide
a)로 전환된다. 또한, 20개의 탄소원자를 포함하는 geranylgeranyl pyrophosphate (GGPP)는 geranylgeranyl reductase (CHLP)에 의해 환원되어 phytyl pyrophosphate (Phytyl-PP)를 형성한다. MV-Chlide
a와 Phytyl-PP는 Chl synthase (CHLG)에 의해 에스테르화되어 Chl
a를 형성한다. RNAi에 의해
CHLG 발현이 감소된 담배 형질전환체에서는 하위 물질인 Chl
a와 Chl
b 모두가 감소되었을 뿐 아니라 엽록소 생합성의 상위 물질인 ALA 축적과 Mg-chelatase의 활성이 줄어들었다. 반면에
CHLG가 과발현된 담배 형질전환체에서는 ALA 축적과 Mg-chelatase 활성이 현저히 증가되었다(
Shalygo et al. 2009).
CHLG의 활성 억제는 전반적인 엽록소 대사 흐름을 피드백 억제한 반면, 과발현은 엽록소 대사 경로의 활성화를 유도하였다. 이러한 결과는
CHLG의 조절이 엽록소 대사 흐름을 규제하는 중요한 요인임을 나타낸다.
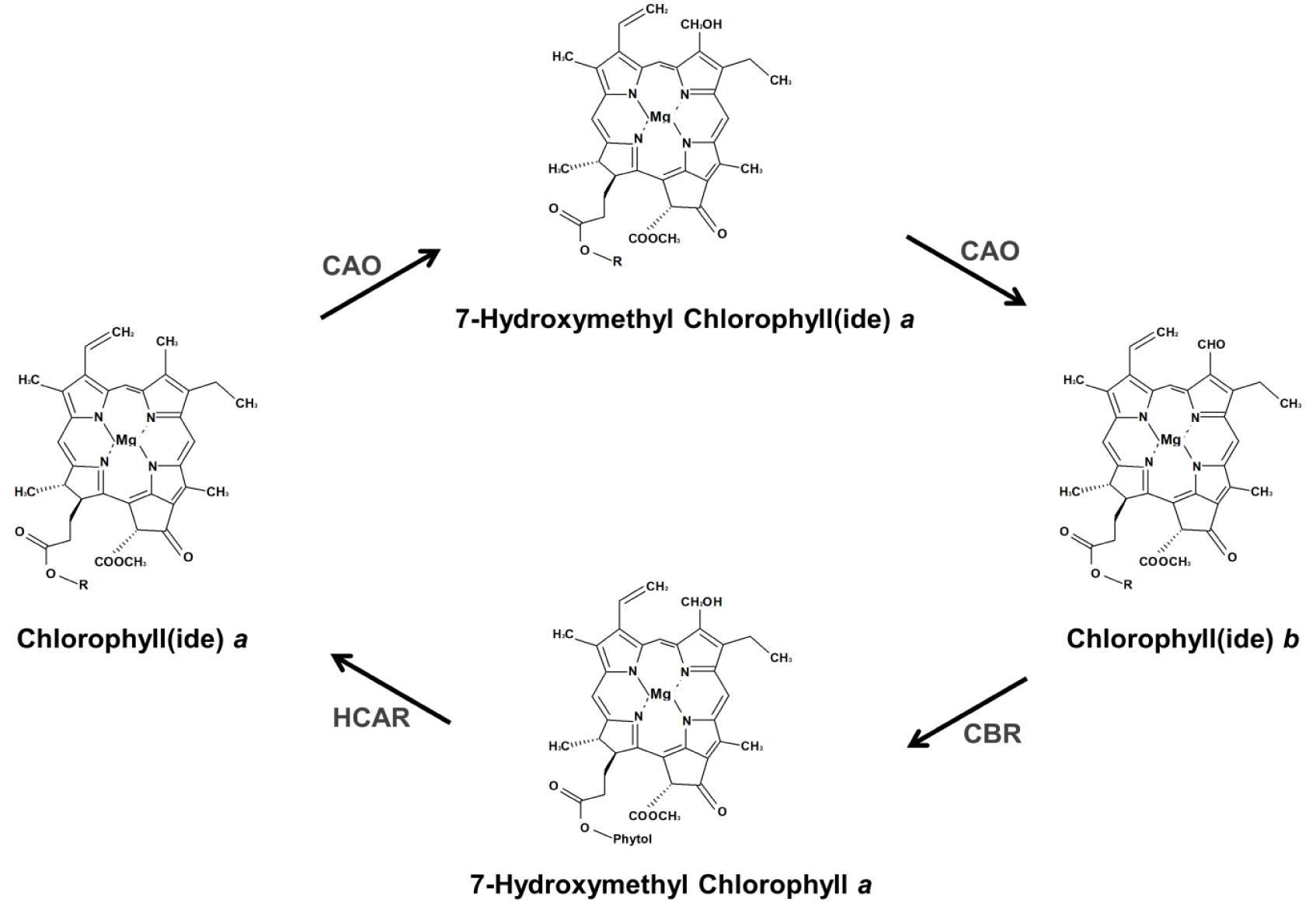
엽록소는 Chl a와 Chl b로 구분되며, Chl a는 광화학 반응에 필수적인 요소인 반면 Chl b는 광합성에 필수적이지 않다. 하지만, Chl b는 식물이 빛에 적응하도록 하며, 엽록소 결합 단백질의 안정화에 필수적이다. Chl a와 b는 흡수하는 빛의 스펙트럼이 다르기 때문에 식물은 넓은 파장의 빛에 반응할 수 있다. Chl b는 Chl a 로부터 합성되고, Chl b reductase (CBR)와 7-hydroxymethyl Chl a reductase (HCAR)에 의해 Chl a로 재전환된다. 이러한 Chl a와 Chl b 사이의 상호전환적 작용을 엽록소 순환이라 하며, 넓은 스펙트럼의 빛을 흡수하기 위한 식물의 조절 기작 중 하나이다.
육상식물의 Chl
b는 광 수확 복합체 결합 단백질의 안정화(stabilization)에 필요하며, Chl
a에서 Chl
b로의 전환이 광 수확 복합체의 생성과 분열을 조절할 수 있다. Chl
a 또는 chlorophyllide
a (Chlide
a)는 Chlide
a oxygenase (CAO)에 의해 hydrocymethyl Chl
a (HMChl
a) 또는 hydrocymethyl Chlide
a (HMChlide
a)로 변환된 후, Chl
b 또는 Chlide
b로 전환된다(
Fig. 2,
Espineda et al. 1999,
Tanaka et al. 1998). 일반적으로 식물의 Chl
a/
b 비율은 3~4로 보고되었으나, CAO 단백질 수준이 증가됨에 따라 Chl
a/
b 비율이 약 3에서 1이하로 급격하게 감소되었다(
Hirashima et al. 2006).
반면에, 식물의 잎이 노화 될 때
CBR과
HCAR 발현이 증가된다. Chl
b는 CBR와 HCAR에 의해 Chl
a로 전환되며 Chl
b의 수준은 감소하게 된다(
Fig. 2,
Ito et al. 1996,
Scheumann et al. 1996;
1998). 식물의 노화과정 동안, 틸라코이드 막에 위치하는 광 수확 복합체II의 분해에 CBR이 필수적인 역할을 한다고 보고되었다(
Horie et al. 2009).
CBR이 결핍된 벼와 애기장대 돌연변이체의 경우, Chl
b의 분해가 감소하며 광 수확 복합체 II가 안정화되어 틸라코이드 막이 거의 분해되지 않는 반면, 대조구 식물체에서는 잎 노화 동안에 틸라코이드 막이 분해되었다(
Scheumann et al. 1999,
Kusaba et al. 2007,
Horie et al. 2009). 애기장대와 벼의
hcar 돌연변이체에서는 HMChl
a의 축적과 함께 녹색 지속(stay-green) 표현형이 나타나지만, 식물의 영양생장 단계에서 심각한 세포 사멸이 확인되었다(
Meguro et al. 2011). 세포 사멸의 원인을 알아보기 위해, 벼의 원형질체에 일시적으로
HCAR의 전구체인 HMChl
a을 처리한 결과 다량의 일중항산소가 생산되는 것이 확인되었다. 반면,
HCAR 과발현 식물체는 대조구 식물체와 비교했을 때 활성산소에 대해 증가된 내성을 나타냈다. 이러한 결과는 노화된 잎에서
HCAR 과발현이 HMChl
a에서 Chl
a로의 전환에 작용함과 더불어 HMChl
a의 과다축적을 방지함으로써 활성 산소로부터 식물을 보호하는 것으로 여겨진다(
Piao et al. 2017).
Pheophytin
a는 Chl
a에서 Mg
2+을 추출하는 Mg-dechelatase 효소 반응에 의해 생성된다. 애기장대에서 Mg-dechelatase를 암호화하는 세 개의
stay-green (
SGR) 유전자가 보고되었다(
Shimoda et al. 2016). SGR1과 SGR2는 Chl
a에 대한 탈금속 활성(dechelating)을 지니며, 대조적으로 SGRL은 Chl
a보다 Chlide
a에 대해 높은 탈금속 활성을 지녀, Chlide
a에서 Mg
2+를 추출하여 pheophorbide
a로 변환시킨다(
Shimoda et al. 2016). 노화과정 동안 애기장대의
SGR1과
SGRL의 발현은 엽록소 분해를 촉진하나, 흥미롭게도
SGR2의 발현은 엽록소 분해를 음성적으로 조절한다(
Sakuraba et al. 2014). 한편, 모든 SGR은 광계 시스템과 광 수확 복합체를 이용한 효소 실험을 통해 SGR이 단일 엽록소 뿐만 아니라 엽록소-단백질 복합체의 엽록소로부터 Mg
2+을 추출하는 것이 확인되었다. 따라서 SGR은 엽록소 분해에 관여할 뿐만 아니라 광계 시스템 분해에도 작용한다(
Shimoda et al. 2016).
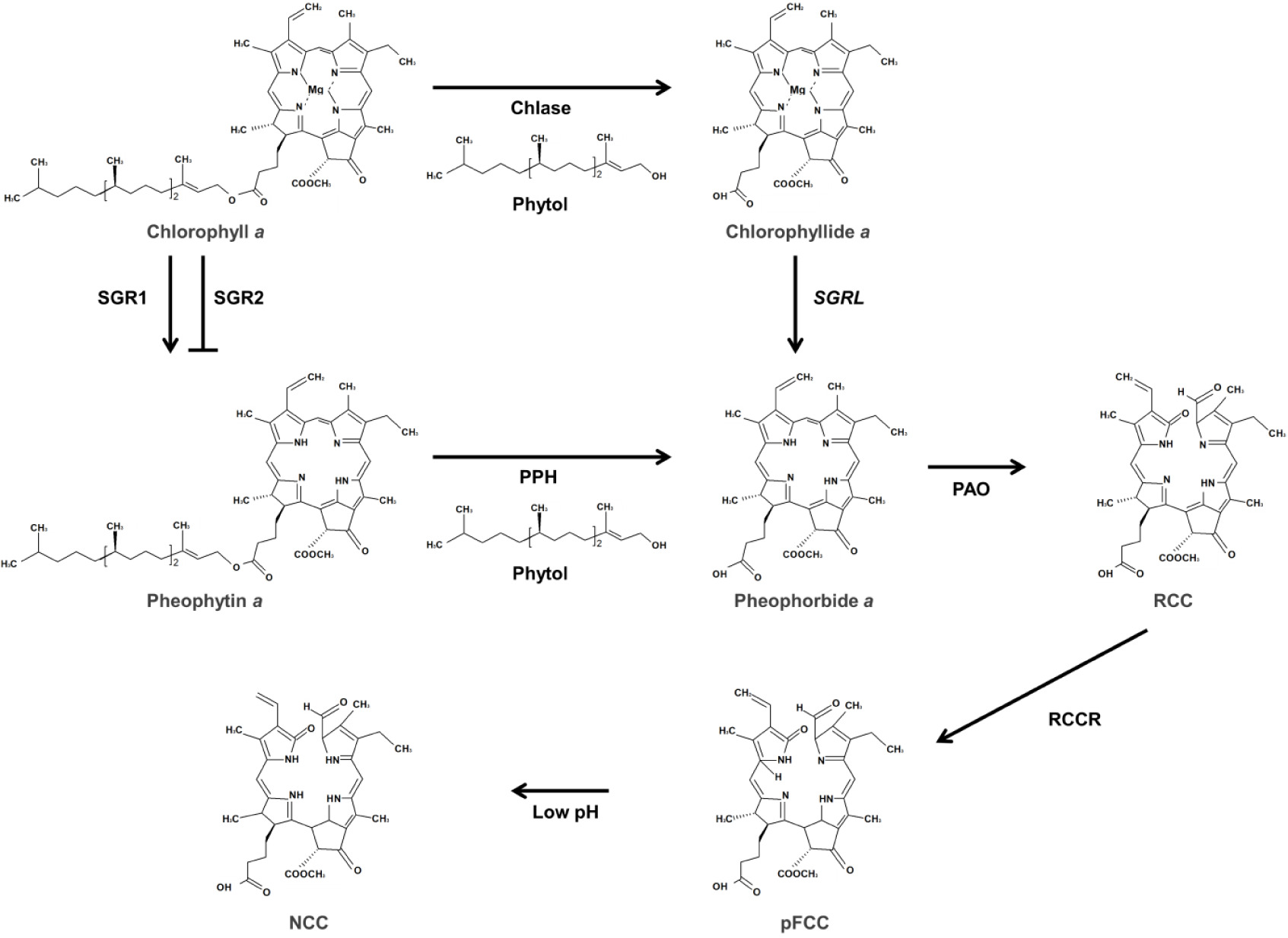
노화 동안 엽록소는 틸라코이드 막에서 제거되고, 여러 단계의 경로를 통해 무색 분해 산물로 전환되어 액포에 저장된다. Pheophytin
a는 pheophytinase (PPH)에 의해 가수분해되어 pheophorbide
a로 변환된다. 애기장대의
pph-1 돌연변이체에서는 노화과정 동안 pheophytin
a가 축적되고 엽록소 분해가 진행되지 않아 잎이 녹색으로 유지되었다. 이러한 결과는 엽록소 분해단계에서 PPH의 중요성을 나타낸다(
Schelbert et al. 2009).
Pheophorbide
a는 pheophorbide
a oxygenase (PAO) 효소에 의해 red Chl catabolite (RCC)로 변환된다. 상시적으로 발현되는 대부분의 엽록소 대사 효소와 달리,
PAO는 노화된 조직에서 국한되어 발현된다. 노화 유도 시, 애기장대의
PAO 유전자의 발현수준은 약간 상향 조절되지만 PAO 단백질 활성이 크게 증가된다(
Pružinská et al. 2003). 이러한 결과를 바탕으로 PAO의 노화특이적 활성은 전사 후 번역 단계에서 조절되어지는 것으로 생각된다.
pFCC는 엽록체로부터 이동되어 비효소적 반응에 의하여 modified FCC (mFCC)로 변환된 후 액포에 축적된다. mFCC는 세포 내의 낮은 pH(4.9)에 반응하여 nonfluorescent Chl catabolite (NCC)로 변환된다(
Fig. 3,
Oberhuber et al. 2003).
전사인자에 의한 엽록소 생합성 조절 기작
엽록소 생합성은 다양한 전사인자군에 의해 조절되어지며, 광 의존적으로 엽록소 생합성 유전자들을 활성화시키는 양성 전사인자와 억제시키는 음성 전사인자로 구분되어지며, 대표적인 전사인자들을
Table 2에 정리하였다.
빛은 식물의 광합성을 위한 에너지원으로 작용하며, 광수용체를 통해 감지된다(
Quail 2002,
Chen et al. 2004). 원적외선과 적색광을 감지하는 광수용체인 피토크롬에 의해 빛을 인지하여 세포내 유전자 발현을 변화시키기 위한 신호전달과정이 시작된다(
Quail 2002,
Chen et al. 2004,
Leivar & Quail 2011). 이때, PIFs와 Long hypocotyl 5 (HY5)는 상반된 방식으로 광반응을 매개한다(
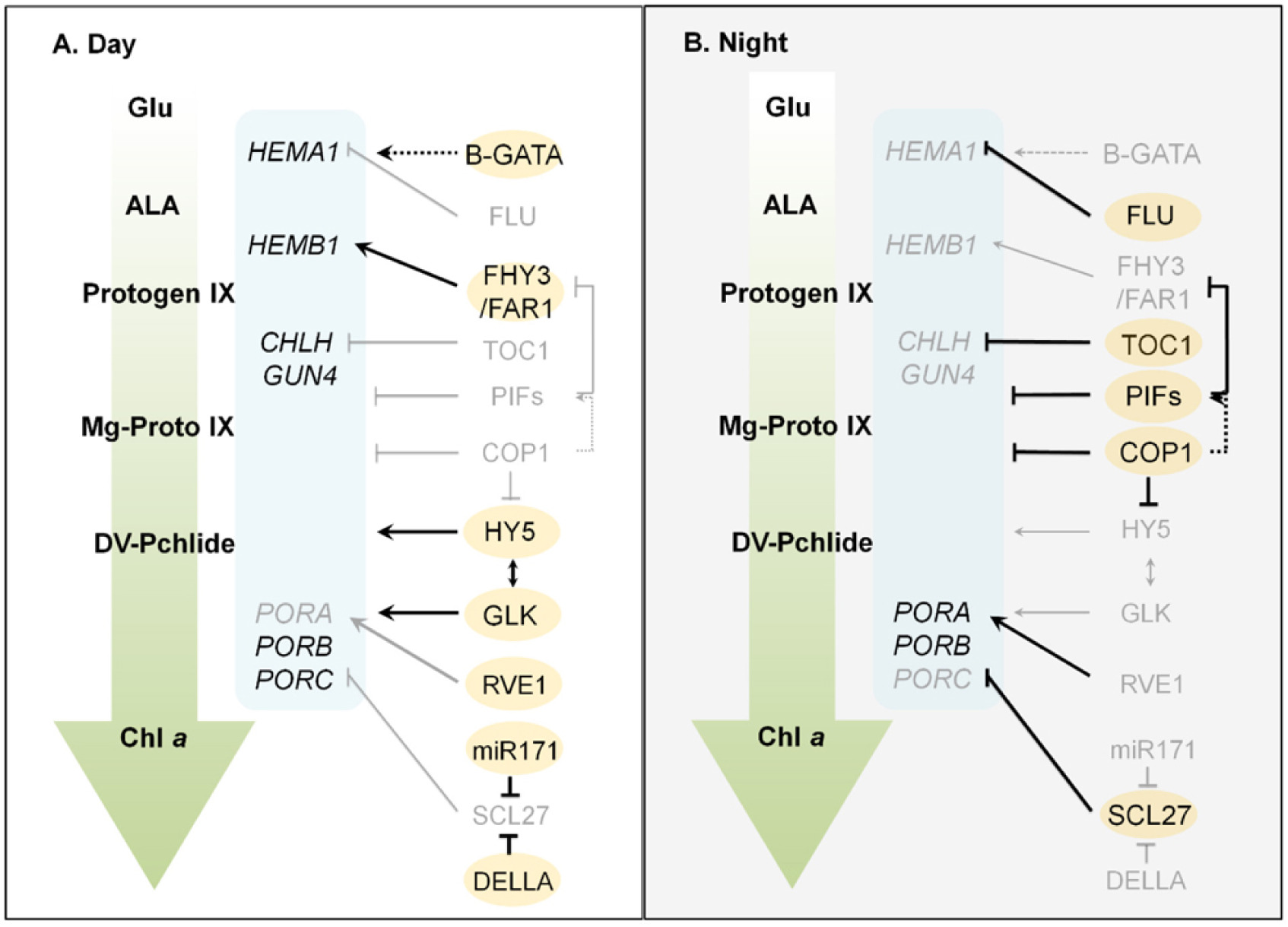
Fig. 4,
von Arnim & Deng 1996,
Chen et al. 2004,
Leivar & Quail 2011). 빛 조건에서 광산화를 유발하는 엽록소 중간 물질의 과도한 축적을 막기 위해, 대다수의 엽록소 생합성 유전자들은 암조건에서 음성적으로 조절된다. 광, 호르몬 및 여러 가지 신호물질에 반응하여 광범위한 세포 발달과정에 관여하는 PIFs 전사인자는 암조건에서 엽록소 생합성 억제에 중요한 역할을 수행한다(
Leivar & Quail 2011). PIFs는 암조건시 핵에 위치하여 엽록소 생합성 및 광형태형성(photomorphogenesis) 관련 유전자들의 발현을 억제한다. 특히, PIF1은 ubiquitin E3 ligase인 constitutive photomorphogenic 1 (COP1)의 보조인자로 작용하여 어둠 속에서 HY5 단백질의 분해를 촉진한다(
Fig. 4,
Zhu et al. 2015). 반면, 빛 조건시 PIFs는 인산화된 후 유비퀴틴에 의한 단백질 분해복합체를 통해 분해되어진다(
Leivar & Quail 2011).
PIF1, PIF3, PIF4 및 PIF5는 주로 암조건에서 유식물이 생장하는 동안 엽록소 생합성 유전자의 발현을 중복적으로 하향 조절한다.
pif1과
pif3 돌연변이는 암상태에서도 다량의 DV-Pchlide를 축적하며, 빛 노출 시에는 심각한 광표백현상이 나타난다(
Huq et al. 2004,
Monte et al. 2004,
Moon et al. 2008,
Shin et al. 2009,
Stephenson et al. 2009). PIF1, PIF3, PIF4 및 PIF5가 결여된 4중 돌연변이(
pifq)에서는 대다수의 광합성 관련 유전자들이 활성화 되어졌다(
Leivar et al. 2009,
Shin et al. 2009). PIFs는 다양한 엽록소 생합성 유전자의 프로모터에 존재하는 G-box (CACGTG)영역에 결합하여 이들의 발현을 억제한다(
Toledo-Ortiz et al. 2014). 특히, PIF1은
PORC의 프로모터에 직접 결합하며, PIF3는
HEMA1,
GUN4 및
CHLH 유전자 발현을 억제한다고 보고되었다(
Moon et al. 2008,
Stephenson et al. 2009).
또한, PIFs는 카로티노이드 생합성의 주요 효소인
phytoene synthase (
PSY) 유전자의 발현을 직접적으로 조절한다(
Toledo-Ortiz et al. 2010). 종자가 발아하여 유식물이 지상으로 올라오는 순간, 암상태에서 빛 상태로의 갑작스런 광환경변화가 발생하게 된다. 빛에 의해 PIFs가 분해되어짐에 따라, 카로티노이드 생합성 및 엽록소 생합성경로 유전자들의 발현이 유도되어지고 카로티노이드계열의 물질 생성도 증가되어진다. 동시에 유식물이 황화과정에서 녹화과정으로 신속하게 변화하면서 광합성이 이루어지고 일중항산소의 축적은 카로티노이드 생합성을 통해 저해된다(
Toledo-Ortiz et al. 2010).
GATA 전사인자는 표적 유전자의 프로모터 영역의 G-A-T-A 코어 서열을 인식하는 조절인자이다. GATA 전사인자군은 식물에 약 30개가 존재하는데, class B에 해당하는 B-GATA는 다양한 식물 발달 과정에서 엽록소 대사를 조절하는 것으로 알려져 있다(
Behringer & Schwechheimer 2015). B-GATAs에 속하는 GNC (GATA, nitrate-inducible, carbon metabolism involved)와 CGA1/GNL (GNC-like/cytokinin-responsive GATA transcription factor 1)은 PIF에 의하여 음성적으로 조절되고 사이토키닌에 의해 유도되며 엽록체 분화 조절에 중요한 역할을 한다.
GNC와
GNL 이중 돌연변이체
gnc gnl에서는 잎에서 엽록소 함량이 감소된 반면,
GNC와
GNL 과발현체에서는 잎 표피, 뿌리, 그리고 하배축에서 엽록소 축적이 증가되었다(
Mara & Irish 2008,
Richter et al. 2010,
Hudson et al. 2011,
Chiang et al. 2012,
Hudson et al. 2013).
gnc gnl 돌연변이체에서
HEMA1,
GUN4,
PORB 및
PORC의 발현은 하향 조절되었지만,
GNC와
GNL의 과발현체에서는 앞서의 유전자들의 발현이 상향 조절되었다(
Hudson et al. 2011). PIF, HY5 및 GLK는 엽록소 생합성 유전자들의 프로모터영역에 결합하여 직접적인 조절 양상을 나타내는 반면, GNC와 GNL은 엽록소 생합성 유전자들의 발현을 간접적으로 조절하는 것으로 생각되어진다(
Fig. 4,
Hudson et al. 2011).
피토크롬A와 상호작용하는 전사인자인 FHY3와 FAR1은 동일한 기능을 하는 상동 단백질로, 엽록소 생합성을 포함하여 다양한 식물 발달 과정에 관여하는 양성 조절인자이다(
Wang & Wang 2015). 애기장대의
fhy3와 far1 돌연변이체에서 근적외선처리가 증가함에 따라
HEMA1,
GUN4, 및
CHLH 발현이 감소됨이 확인되었다(
McCormac & Terry 2002,
Stephenson & Terry 2008). FHY3와 FAR1은 표적 유전자의 프로모터영역에 존재하는 FHY3/FAR1 결합 부위(CACGCGC)에 결합한다. 특히, FHY3/FAR1은 ALAD를 암호화하는
HEMB1의 프로모터에 직접적으로 결합하며, 그 중에서 FHY3가
HEMB1을 우세하게 조절함이 보고되었다(
Ouyang et al. 2011,
Tang et al. 2012). 또한, PIF1과 FHY3는 물리적으로 상호 결합하며, 암조건시 FHY3는 PIF1에 의해 억제되어
HEMB1의 발현을 유도하지 못하여 Pchlide의 축적이 감소된다(
Fig. 4,
Tang et al. 2012). 이러한 결과는 FHY3/FAR1이
HEMA1,
GUN4, 및
CHLH 발현에도 영향을 줄 뿐만 아니라,
HEMB1을 직접적으로 조절하는 것을 나타낸다(
McCormac & Terry 2002,
Stephenson & Terry 2008).
MicroRNA miR171과 그의 표적 전사인자인 SCLs (SCL6/22/27)은 엽록소 특이적 생합성 경로의 전사 조절에 중요한 역할을 한다(
Wang et al. 2010,
Ma et al. 2014). miR171의 과발현은 표적 SCLs 전사체를 분해시켰으며,
POR를 상향 조절하여 엽록소 축적을 증가시켰다. 이와 대조적으로,
SCLs 중
SCL27의 과발현은 잎에서 엽록소 축적을 감소시켰다. 생체 내와 시험관 내 분석을 통하여, SCL27이
PORC 프로모터 내 GT
cis-element [G(A/G)(A/T)AA(A/T)]에 결합하여 유전자의 발현을 억제함이 보고되었다(
Ma et al. 2014). 따라서 miR171과 이들의 표적
SCLs 사이의 균형은 잎에서 엽록소 생합성 경로 유전자들의 발현에 영향을 준다. 애기장대의
ga1-3 돌연변이체에서 지베렐린 신호 전달의 음성조절자인 DELLA 단백질이
SCLs의 전사활성을 억제하여 엽록소 생합성을 증가시키는 것이 확인되었다(
Ma et al. 2014). 이러한 결과는 엽록소 생합성 유전자들의 발현 조절이 지베렐린 신호 전달과도 연관성이 있으며, SCLs이 엽록소 생합성에 음성조절자로 작용한다는 것을 확인할 수 있다(
Ma et al. 2014).
RVE 전사인자는 evening
cis-element (AAAATATCT)에 결합하여 암조건에서
PORA 발현을 상향 조절한다(
Fig. 4,
Xu et al. 2015).
RVE1이 과발현된 애기장대에서는 암조건에서 활성산소 생성과 세포 사멸이 감소되었고, 유식물의 녹화 비율은 증가되었다. 이는 RVE1이
PORA 발현을 직접적으로 증가시켜 Pchlide의 축적의 감소되었기 때문이다. 따라서, RVE1은 엽록소 생합성을 조절하며, 식물의 성장과 발달 동안 유식물의 녹화를 촉진한다(
Xu et al. 2015).
결 론
엽록소는 광합성의 광 수확 및 에너지 변환에 필수적인 역할을 수행하는 식물 색소로, 광합성 기관인 엽록체에서 생합성이 이루어진다. 생화학적 및 분자생물학적 분석을 통해 엽록소 생합성과 분해에 관여하는 유전자들이 여러 식물에서 동정 되었다. 엽록소 생합성 유전자들의 발현은 빛 조건과 일주기 신호에 의해서 조절되어지는 것으로 보고되고 있다. 일반적으로 빛 조건에서 엽록소 생합성 관련 유전자들의 발현이 상향 조절되어지며 암 조건에서는 엽록소 분해와 관련된 유전자들의 발현이 증대되어지는데, 이러한 엽록소 생합성 및 분해 관련 유전자들의 발현은 광합성 관련 유전자들과 밀접하게 공동 발현되어짐이 확인되었다. 또한 여러 연구자들은 프로모터의 결합 부위 분석을 통해 엽록소 생합성 관련 유전자들을 공동 조절하는 양성 혹은 음성 조절인자들을 동정하고 그 기능을 확인하여 엽록소 생성 및 분해 기작을 포괄적으로 이해하고자 시도하고 있다. 돌연변이체 및 유전자 과발현을 통한 여러 연구를 통해서 엽록소 생합성 과정 중 중간 물질의 축적은 광산화를 발생시켜 식물 생장에 부정적인 영향을 주는 것이 확인되었다. 따라서 식물체의 엽록소 생성은 대사채널링을 통해 중간 물질의 방출을 최소화하는 방식으로 정교하게 조절되어진다고 생각된다.
여러 연구를 바탕으로 식물체 내의 엽록소의 함량은 생합성과 분해 간의 상호 조절을 통해서 이루어짐이 확인되었다. 일반적으로 녹화과정에서 생합성 유전자의 높은 발현 수준과 단백질의 안정화는 엽록소의 축적을 유도하여 광합성을 촉진시킨다. 반면, 노화과정 동안에는 분해과정관련 유전자들의 발현이 상대적으로 증가됨에 따라 엽록소의 분해는 촉진되어진다. 최근 다수의 연구를 통하여 광합성에 필수적인 식물 색소인 엽록소의 축적은 다양한 외부환경(온도 및 호르몬)에 의해 조절됨과 더불어 식물체가 다양한 외부 불량 환경에 적응하는 것에 도움을 주는 것이 밝혀졌다.
엽록소 생성 및 분해에 관한 연구의 중요성에도 불구하고, 현재까지는 몇몇 모델식물에 국한되어 연구가 진행되었다. 본 논문에서 보고한 엽록소 생성 및 분해에 관한 연구결과들은 다양한 작물의 엽록소 생합성 기작을 이해하는데 도움을 줄 수 있을 것이다. 또한 엽록소는 광합성에 매우 중요한 잎 뿐 아니라 꽃과 과실과 같은 다양한 조직에서도 축적되기 때문에, 엽록소 생합성 기작에 대한 이해는 다양한 작물에서 엽록소 축적에 대한 과학적인 고찰을 제공하며, 새로운 육종 소재 개발에도 도움이 되리라 예상된다.
적 요
엽록소는 식물 생장과 발달에 매우 중요한 광합성 작용에서 빛의 흡수와 전자 전달에 관여하는 필수적인 색소물질이다. 엽록소의 생합성은 색소체 내에서 일어나며, 다른 테트라피롤계 물질들과 일부 경로를 공유한다. 엽록소 경로는 크게 공통 경로, 엽록소 특이적 경로, 엽록소 순환, 그리고 엽록소 분해로 네 단계로 나뉘며 식물의 생장 및 환경에 따라 각각의 경로가 조절되어진다. 식물의 생장초기에는 빛에 의해 대부분의 엽록소 생합성 유전자들의 발현이 유도되어 엽록소 생합성이 증가하고, 높은 광합성 활성과 지속적인 식물 성장이 유도된다. 대조적으로 식물 성숙기에 대다수의 엽록소 생합성 유전자들의 발현이 점차적으로 감소되며, 잎 노화기에는 거의 발현되지 않으며 분해 경로에 해당하는 유전자들의 발현이 증대된다. 엽록소 생합성은 다양한 전사인자군에 의해 직간접적으로 조절되어지며, 본문에는 광에 의해 엽록소 생성을 활성 혹은 억제하는 대표적인 전사인자들의 작용 기작을 기술하였다. 본 리뷰는 주요 모델 작물인 애기장대에서 밝혀진 유전자들의 기능에 대하여 서술하였으며, 이를 통하여 엽록소 생합성과 전사 조절에 대한 전반적인 이해를 돕고자 하였다.
사 사
본 연구는 농촌진흥청 국립농업과학원의 공동연구과제(과제명: 무 유전자원의 청피, 백피 구분 마커 개발, 과제번호: PJ014554012020)의 지원에 의해 이루어진 것으로 이에 감사드립니다.
Fig. 1Schematic representation of chlorophyll metabolic pathways in Arabidopsis. The dotted grey lines represent the individual four distinct pathways. The abbreviations of enzyme names and metabolites are as follows: 1) Common pathway; GluRS, glutamyl-tRNA synthetase; GluTR, glutamate-tRNA reductase; GSA, glutamate 1-semialdehyde aminotransferase; ALAD, 5-aminolevulinic acid dehydratase; PBGD, porphobilinogen deaminase; UROS, uroporphyrinogen III synthase; UROD, uroporphyrinogen III decarboxylase; CPO, coprophyrinogen III oxidase; Glu, glutamate; GSA, Glu 1-semialdehyde; ALA, 5-aminolevulinic acid; PBG, monopyrrole porphobilinogen; HMB, hydrocymethylbilane; Urogen III, uroporphyrinogen III; Coprogen III, coprophyrinogen III; FC, ferrochelatase. 2) Chlorophyll specific biosynthetic pathway; PPO, protoporphyrinogen IX oxidase; CHLD/CHLH/CHLI, Mg-chelatase subunit D/H/I; GUN4, genomes uncoupled 4; CHLM, Mg-protoporphyrin IX methyltransferase; CHL27/LCAA, low chlorophyll accumulation A, Mg-protoporphyrin IX monomethylester cyclase subunit; Mg-protoporphyrin IX monomethyl ester cyclase; POR, NADPH:protochlorophyllide oxidoreductase; DVR, 3,8-divinyl-chlorophyllide reductase; CHLP, geranylgeranyl reductase; CHLG, chlorophyll synthase; Protogen IX, protoporphyrinogen IX; Proto IX, protoporphyrin IX; Mg-Proto IX, Mg-protoporphyrin IX; Mg-ProtoME, Mg-protoporphyrin IX monomethylester; DV-Pchlide, protochlorophyllide; DV-Chlide, 3,8-divinyl-chlorophyllide; MV Chlide a, 3-vinyl chlide a; GGPP, geranylgeranyl pyrophosphate; Phytyl-PP, phytyl pyrophosphate. 3) Chlorophyll cycle pathway; Chl(ide) a, chlorophyll(ide) a; HMChl (ide) a, 7-hydrocymethyl chlorophyll(ide) a; Chl(ide) b, chlorophyll (ide) b; HMChl a, 7-hydrocymethyl chlorophyll a. 4) Chlorophyll degradation pathway; RCC, red chlorophyll catabolite; pFCC, primary fluorescent chlorophyll catabolites; mFCC, modified primary fluorescent chlorophyll catabolites; NCC, nonfluorescent chlorophyll catabolite.
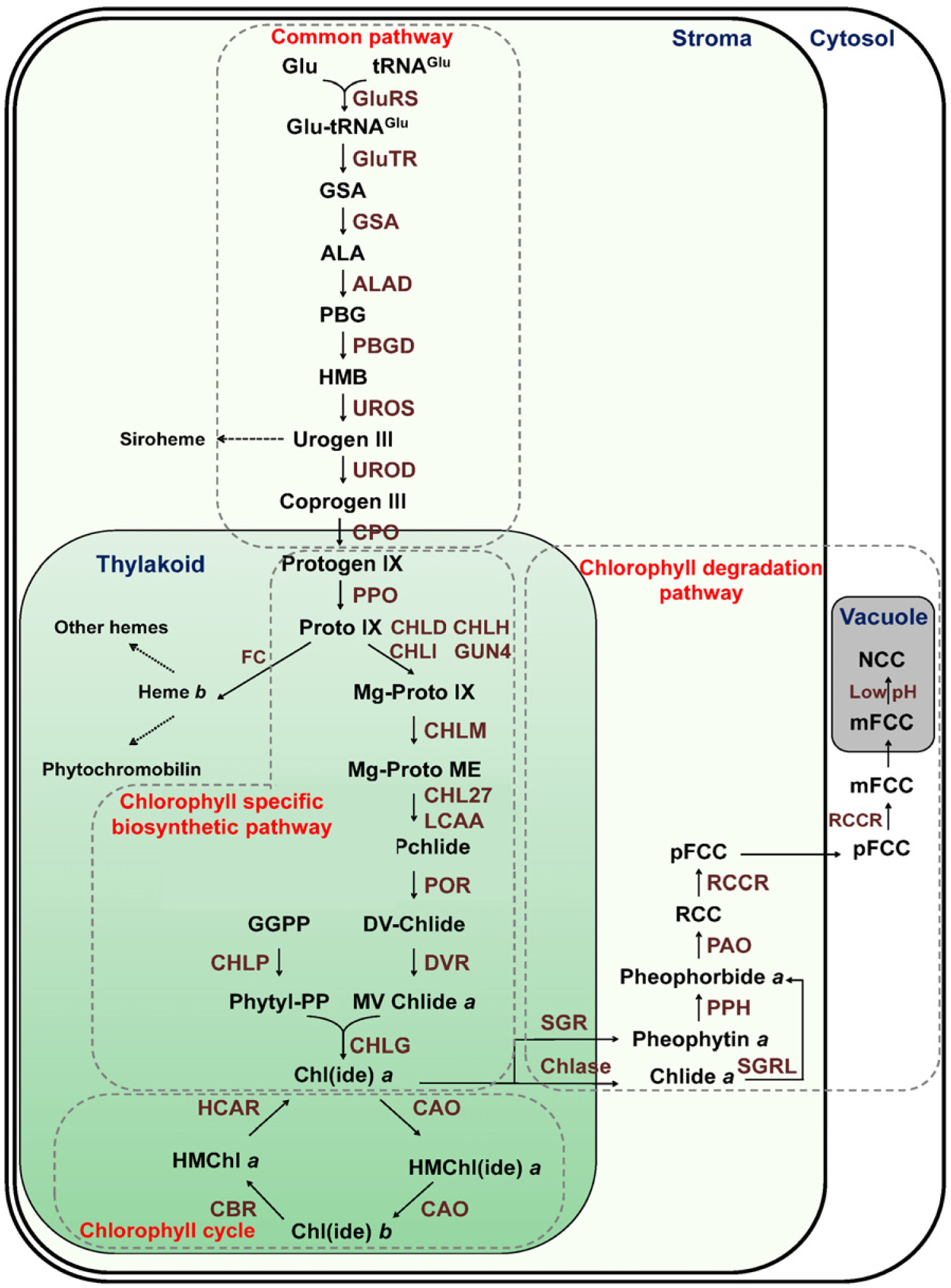
Fig. 2Schematic diagram of chlorophyll cycle. R indicates either protons or phytols. CAO, chlorophyllide a oxygenase; CBR, chlorophyll b reductase; HCAR, 7-hydroxymethyl chlorophyll a reductase.
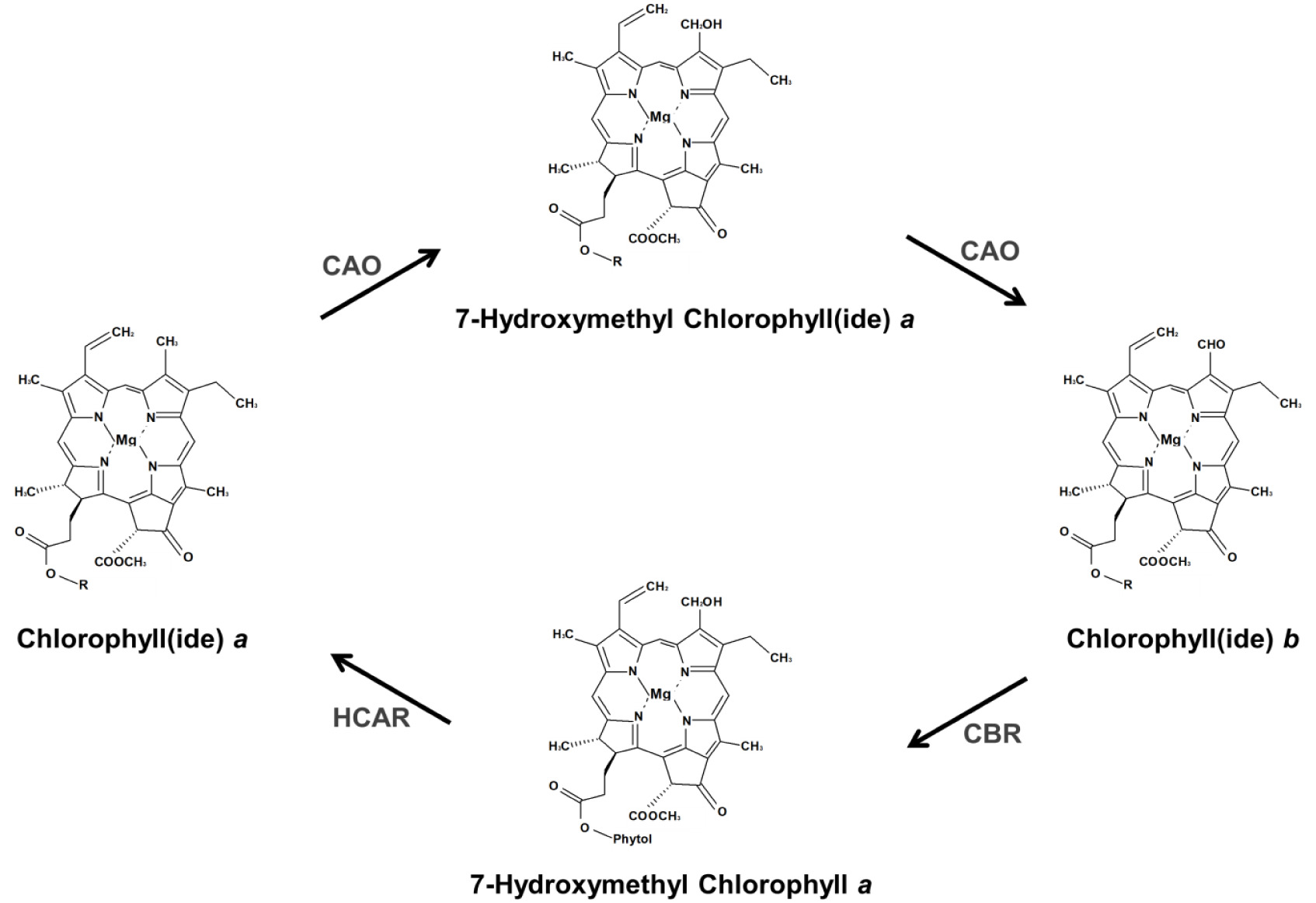
Fig. 3Proposed route of chlorophyll degradation. Chlorophyll a is hydrolyzed by chlase to from chlorophyllide and phytol. Also, chlorophyll is released of Mg by Mg-dechelatase. Chlase, chlorophyllase; SGR, stay-green; PPH, pheophytinase; PAO, pheophorbide a oxygenase; RCCR, red chlorophyll catabolite reductase; pFCC, primary fluorescent chlorophyll catabolites; NCC, nonfluorescent chlorophyll catabolite.
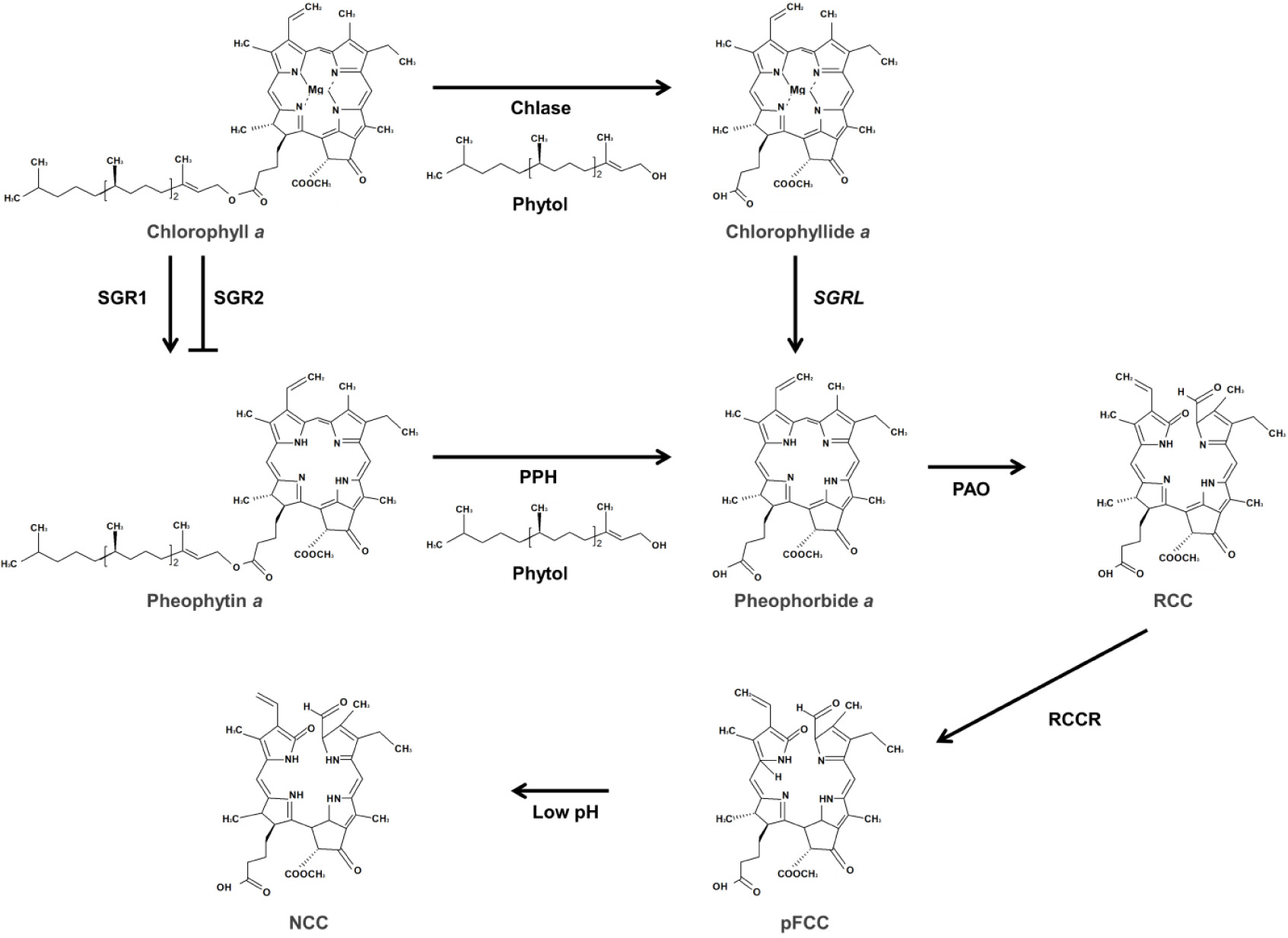
Fig. 4Chlorophyll biosynthetic pathway controlled by transcription factors in day (A) and night (B). Bold letters and lines represent activated status. Solid arrows and solid bars indicate the positive and negative regulation, respectively. Dotted lines indicate indirect effects (
Behringer & Schwechheimer 2015,
Chattopadhyay et al. 1998,
Goslings et al. 2004,
Leivar & Quail 2011,
Ma et al. 2014,
Matsumoto et al. 2004,
Tamai et al. 2002,
Tang et al. 2012,
Waters et al. 2009,
Xu et al. 2015,
Zhu et al. 2015).
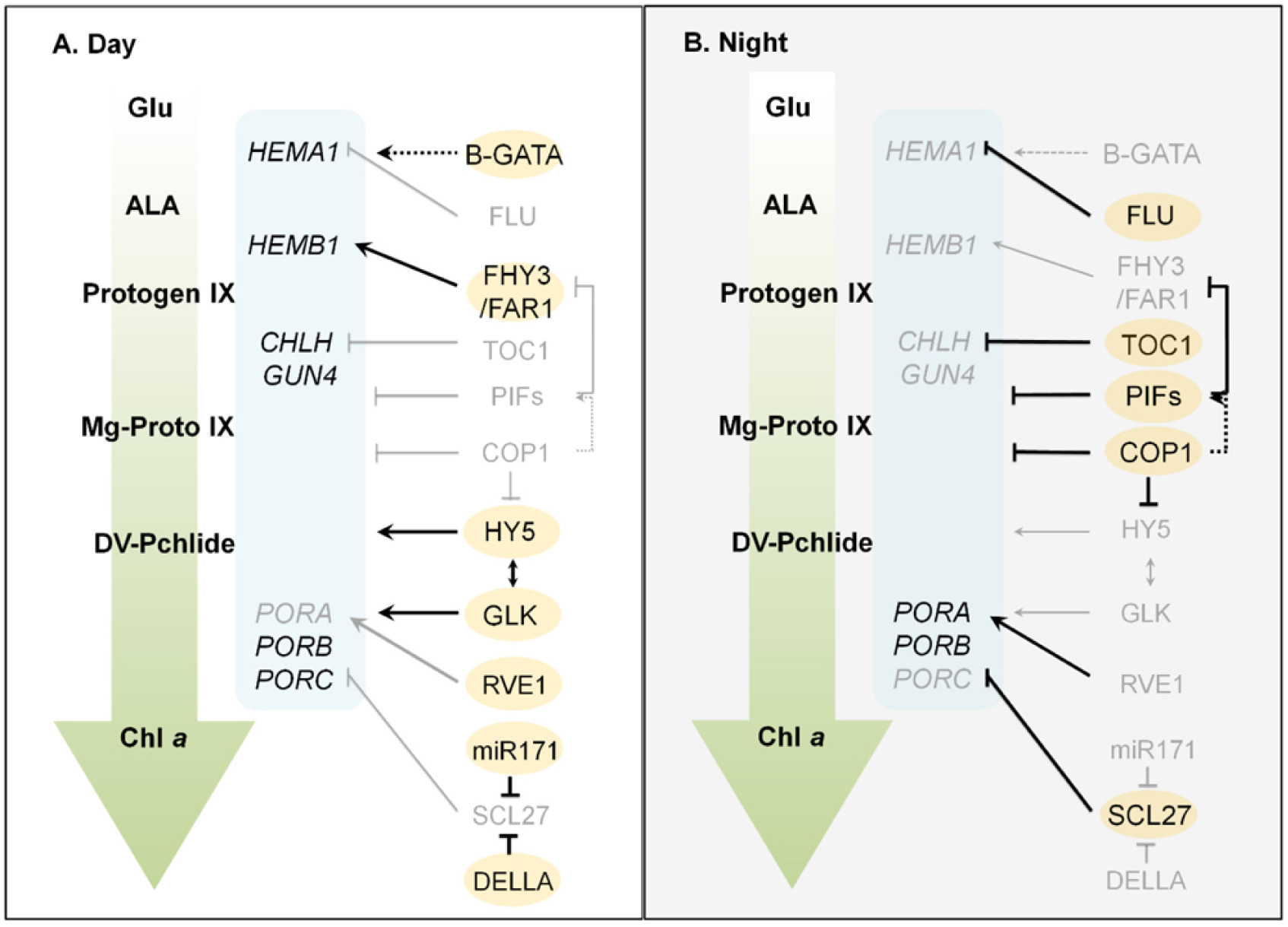
Table 1Enzymes involved in chlorophyll biosynthetic pathway.
Table 1
|
Metabolic pathway |
Enzyme name |
Arabidopsis |
Rice |
Reference |
|
Common pathway |
GluTR |
At1g58290 |
Os10g0502400 |
McCormac et al. 2001, Zheng et al. 2012 |
|
At1g09940 |
|
At2g31250 |
|
GSA1 |
At5g63570 |
Os08g0532200 |
Dalal & Tripathy 2012, Ilag et al. 1994
|
|
GSA2 |
At3g48730 |
|
ALAD |
At1g69740 |
Os06g0704600 |
Dalal & Tripathy 2012, Tang et al. 2012
|
|
At1g44318 |
|
PBGD |
At5g08280 |
Os02g0168800 |
Lim et al. 1994, Parker et al. 2006
|
|
UROS |
At2g26540 |
Os03g0186100 |
Li et al. 2017, Tan et al. 2008
|
|
UROD1 |
At3g14930 |
Os01g0622300 |
Tanaka et al. 2011, Zheng et al. 2013 |
|
UROD2 |
At2g40490 |
Os03g0337600 |
|
CPO1 |
At1g03475 |
Os04g0610800 |
Ishikawa et al. 2001, Matsumoto et al. 2004, Sun et al. 2011
|
|
CPO2 |
At4g03205 |
Chlorophyll specific
biosynthetic pathway |
PPO1 |
At4g01690 |
Os01g0286600 |
Park et al. 2012, Phung et al. 2011, Zhang et al. 2014
|
|
PPO2 |
At5g14220 |
Os04g0490100 |
|
CHLD |
At1g08520 |
Os03g0811100 |
Kobayashi et al. 2008, Zhang et al. 2016 |
|
CHLH |
At5g13630 |
Os03g0323200 |
Adhikari et al. 2011, Jung et al. 2003
|
|
CHLI1 |
At4g18480 |
Os03g0563300 |
Kobayashi et al. 2008, Zhang et al. 2016 |
|
CHLI2 |
At5g45930 |
|
GUN4 |
At3g59400 |
Os11g0267000 |
Li et al. 2014, Richter et al. 2016
|
|
CHLM |
At4g25080 |
Os06g0132400 |
Richter et al. 2016, Wang et al. 2017
|
|
CHL27 |
At3g56940 |
Os01g0279100 |
Rzeznicka et al. 2005, Wang et al. 2017
|
|
LCAA |
At5g58250 |
Os03g0331600 |
Albus et al. 2012, Kong et al. 2016
|
|
PORA |
At5g54190 |
Os04g0678700 |
Eckhardt et al. 2004, Sakuraba et al. 2013
|
|
PORB |
At4g27440 |
Os10g0496900 |
|
PORC |
At1g03630 |
|
DVR |
At5g18660 |
Os03g0351200 |
Eckhardt et al. 2004, Wang et al. 2010
|
|
CHLP |
At1g74470 |
Os01g0265000 |
Kimura et al. 2018, Takahashi et al. 2014
|
|
CHLG |
At3g51820 |
Os05g0349700 |
Lin et al. 2014, Zhou et al. 2017
|
|
Chlorophyll cycle |
CAO |
At1g44446 |
Os10g0567100 |
Espineda et al. 1999, Lee et al. 2005
|
|
CBR |
At5g17770 |
Os05g0488900 |
Horie et al. 2009, Sato et al. 2009
|
|
HCAR |
At1g04620 |
Os04g0320100 |
Meguro et al. 2011, Piao et al. 2017
|
Chlorophyll
degradation
pathway |
Chlase1 |
At1g19670 |
Os10g0419600 |
Benedetti & Arruda 2002, Sato et al. 2007
|
|
Chlase2 |
At5g43860 |
|
SGR1 |
At4g22920 |
Os09g0532000 |
Jiang et al. 2007, Rong et al. 2013, Shimoda et al. 2016
|
|
SGR2 |
At4g11910 |
|
SGRL |
At1g44000 |
Os04g0692600 |
|
PPH |
At5g13800 |
Os06g0354700 |
Morita et al. 2009, Schelbert et al. 2009
|
|
PAO |
At3g44880 |
Os03g0146400 |
Pružinská et al. 2003, Tang et al. 2011
|
|
RCCR |
At4g37000 |
Os10g0389300 |
Pružinská et al. 2007, Tang et al. 2011
|
Table 2Transcription factors related on chlorophyll biosynthetic pathway.
Table 2
|
Regulatory pattern |
Transcription factor |
Arabidopsis |
Rice |
Reference |
|
Positive |
HY5 |
At5g11260 |
Os06g0601500 |
Burman et al. 2018, Toledo-Ortiz et al. 2014
|
|
Positive |
GLK1 |
At2g20570 |
Os06g0348800 |
Nakamura et al. 2009, Wang et al. 2013, Waters et al. 2009
|
|
GLK2 |
At5g44190 |
Os01g0239000 |
|
Positive |
GNC |
At5g56860 |
Os06g0571800 |
Hudson et al. 2011, Kusano et al. 2020
|
|
Positive |
CGA1/GNL |
At4g26150 |
|
Positive |
FHY3 |
At3g22170 |
Os03g0181600 |
Reyes et al. 2004, Tang et al. 2012
|
|
Positive |
FAR1 |
At4g15090 |
Os07g0615900 |
|
Positive |
RVE |
At5g17300 |
Os02g0685200 |
Gray et al. 2017, Xu et al. 2015
|
|
Negative |
PIF1 |
At2g20180 |
OS04g6618600 |
Li et al. 2016, Zhu et al. 2015
|
|
PIF3 |
At1g09530 |
Os01g0286100 |
Leivar & Quail 2011, Zhou et al. 2013 |
|
PIF4 |
At2g43010 |
Os03g0782500 |
Leivar et al. 2009, Piao et al. 2015
|
|
PIF5 |
At3g59060 |
Os12g0610200 |
Leivar et al. 2009, Nakamura et al. 2007
|
|
Negative |
FLU |
At3g14110 |
Os01g0510600 |
Kong et al. 2016, Meskauskiene et al. 2001
|
|
Negative |
SCL27 |
At2g45160 |
- |
Ma et al. 2014
|
|
Negative |
TOC1 |
At5g61380 |
Os02g0618200 |
Legnaioli et al. 2009, Murakami et al. 2007
|
References
- 1. Adhikari ND, Froehlich JE, Strand DD, Buck SM, Kramer DM, Larkin RM. 2011. GUN4-porphyrin complexes bind the ChlH/GUN5 subunit of Mg-chelatase and promote chlorophyll biosynthesis in Arabidopsis. Plant Cell 23: 1449-1467.
- 2. Albus CA, Salinas A, Czarnecki O, Kahlau S, Rothbart M, Thiele W, Schöttler MA. 2012. LCAA, a novel factor required for magnesium protoporphyrin monomethylester cyclase accumulation and feedback control of aminolevulinic acid biosynthesis in tobacco. Plant Physiol 160: 1923-1939.
- 3. Apitz J, Schmied J, Lehmann MJ, Hedtke B, Grimm B. 2014. GluTR2 complements a hema1 mutant lacking glutamyl-tRNA reductase 1, but is differently regulated at the post-translational level. Plant Cell Physiol 55: 645-657.
- 4. Bae G, Choi G. 2008. Decoding of light signals by plant phytochromes and their interacting proteins. Plant Biol 59: 281-311.
- 5. Behringer C, Schwechheimer C. 2015. B-GATA transcription factors - insights into their structure, regulation, and role in plant development. Front Plant Sci 6: 90
- 6. Benedetti CE, Arruda P. 2002. Altering the expression of the chlorophyllase gene ATHCOR1 in transgenic Arabidopsis caused changes in the chlorophyll-to-chlorophyllide ratio. Plant Physiol 128: 1255-1263.
- 7. Brzezowski P, Richter AS, Grimm B. 2015. Regulation and function of tetrapyrrole biosynthesis in plants and algae. Biochim Biophys Acta 1847: 968-985.
- 8. Burman N, Bhatnagar A, Khurana JP. 2018. OsbZIP48, a HY5 transcription factor ortholog, exerts pleiotropic effects in light-regulated development. Plant Physiol 176: 1262-1285.
- 9. Chattopadhyay S, Puente P, Deng XW, Wei N. 1998. Combinatorial interaction of light-responsive elements plays a critical role in determining the response characteristics of light-regulated promoters in Arabidopsis. Plant J 15: 69-77.
- 10. Chen M, Chory J, Fankhauser C. 2004. Light signal transduction in higher plants. Annu Rev Genet 38: 87-117.
- 11. Chiang YH, Zubo YO, Tapken W, Kim HJ, Lavanway AM, Howard L. 2012. Functional characterization of the GATA transcription factors GNC and CGA1 reveals their key role in chloroplast development, growth, and division in Arabidopsis. Plant Physiol 160: 332-348.
- 12. Chow KS, Singh DP, Walker AR, Smith AG. 1998. Two different genes encode ferrochelatase in Arabidopsis: mapping, expression and subcellular targeting of the precursor proteins. Plant J 15: 531-541.
- 13. Dalal VK, Tripathy BC. 2012. Modulation of chlorophyll biosynthesis by water stress in rice seedlings during chloroplast biogenesis. Plant Cell Environ 35: 1685-1703.
- 14. Davison PA, Schubert HL, Reid JD, Iorg CD, Heroux A, Hill CP, Hunter CN. 2005. Structural and biochemical characterization of Gun4 suggests a mechanism for its role in chlorophyll biosynthesis. Biochem 44: 7603-7612.
- 15. Eckhardt U, Grimm B, Hörtensteiner S. 2004. Recent advances in chlorophyll biosynthesis and breakdown in higher plants. Plant Mol Biology 56: 1-14.
- 16. Espineda CE, Linford AS, Devine D, Brusslan JA. 1999. The AtCAO gene, encoding chlorophyll a oxygenase, is required for chlorophyll b synthesis in Arabidopsis thaliana. Proc Natl Acad Sci USA 96: 10507-10511.
- 17. Fitter DW, Martin DJ, Copley MJ, Scotland RW, Langdale JA. 2002. GLK gene pairs regulate chloroplast development in diverse plant species. Plant J 31: 713-727.
- 18. Gendron JM, Pruneda-Paz JL, Doherty CJ, Gross AM, Kang SE, Kay SA. 2012. Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proc Natl Acad Sci USA 109: 3167-3172.
- 19. Goslings D, Meskauskiene R, Kim C, Lee KP, Nater M, Apel K. 2004. Concurrent interactions of heme and FLU with Glu tRNA reductase (HEMA1), the target of metabolic feedback inhibition of tetrapyrrole biosynthesis, in dark- and light-grown Arabidopsis plants. Plant J 40: 957-967.
- 20. Gray JA, Shalit-Kaneh A, Chu DN, Hsu PY, Harmer SL. 2017. The REVEILLE clock genes inhibit growth of juvenile and adult plants by control of cell size. Plant Physiol 173: 2308-2322.
- 21. Hirashima M, Satoh S, Tanaka R, Tanaka A. 2006. Pigment shuffling in antenna systems achieved by expressing prokaryotic chlorophyllide a oxygenase in Arabidopsis. J Biol Chem 281: 15385-15393.
- 22. Hollingshead S, Kopeccná a J, Jackson PJ, Canniffe DP, Davison PA, Dickman MJ, Sobotka R, Hunter CN. 2012. Conserved chloroplast open-reading frame ycf54 is required for activity of the magnesium protoporphyrin monomethylester oxidative cyclase in Synechocystis PCC 6803. J Biol Chem 287: 27823-27833.
- 23. Holm M, Ma L, Qu LJ, Deng XW. 2002. Two interacting bZIP proteins are direct targets of COP1-mediated control of light-dependent gene expression in Arabidopsis. Genes Dev 16: 1247-1259.
- 24. Horie Y, Ito H, Kusaba M, Tanaka R, Tanaka A. 2009. Participation of chlorophyll b reductase in the initial step of the degradation of light-harvesting chlorophyll a/b-protein complexes in Arabidopsis. J Biol Chem 284: 17449-17456.
- 25. Hudson D, Guevara D, Yaish MW, Hannam C, Long N, Clarke JD, Bi YM, Rothstein SJ. 2011. GNC and CGA1 modulate chlorophyll biosynthesis and glutamate synthase (GLU1/Fd-GOGAT) expressionin Arabidopsis. PLoS One 6: e26765
- 26. Hudson D, Guevara DR, Hand AJ, Xu Z, Hao L, Chen X, Zhu T, Bi YM, Rothstein SJ. 2013. Rice cytokinin GATA transcription factor1 regulates chloroplast development and plant architecture. Plant Physiol 162: 132-144.
- 27. Huq E, Al-Sady B, Hudson M, Kim C, Apel K, Quail PH. 2004. Phytochrome-interacting factor 1 is a critical bHLH regulator of chlorophyll biosynthesis. Science 1937-1941.
- 28. Ilag LL, Kumar AM, Söll D. 1994. Light regulation of chlorophyll biosynthesis at the level of 5-aminolevulinate formation in Arabidopsis. Plant Cell 6: 265-275.
- 29. Ishikawa A, Okamoto H, Iwasaki Y, Asahi T. 2001. A deficiency of coproporphyrinogen III oxidase causes lesion formation in Arabidopsis. Plant J 27: 89-99.
- 30. Ito H, Ohtsuka T, Tanaka A. 1996. Conversion of chlorophyll b to chlorophyll a via 7-hydroxymethyl chlorophyll. J Biol Chem 271: 1475-1479.
- 31. Jakob-Wilk D, Holland D, Goldschmidt EE, Riov J, Eyal Y. 1999. Chlorophyll breakdown by chlorophyllase: isolation and functional expression of the Chlase1 gene from ethylene-treated Citrus fruit and its regulation during development. Plant J 20: 653-661.
- 32. Jiang H, Li M, Liang N, Yan H, Wei Y, Xu X, Liu J, Xu Z, Chen F, Wu G. 2007. Molecular cloning and function analysis of the stay green gene in rice. Plant J 52: 197-209.
- 33. Jung KH, Hur J, Ryu CH, Choi Y, Chung YY, Miyao A, Hirochika H, An G. 2003. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol 44: 463-472.
- 34. Kim JH, Lee MH, Moon YR, Kim JS, Wi SG, Kim TH, Chung BY. 2009. Characterization of metabolic disturbances closely linked to the delayed senescence of Arabidopsis leaves after γ irradiation. J Exp Bot 67: 363-371.
- 35. Kimura E, Abe T, Murata K, Kimura T, Otoki Y, Yoshida T, Miyazawa T, Nakagawa K. 2018. Identification of OsGGR2, a second geranylgeranyl reductase involved in α-tocopherol synthesis in rice. Sci Rep 8: 1-8.
- 36. Kobayashi K, Baba S, Obayashi T, Sato M, Toyooka K, Keränen M, Aro EM, Fukaki H, Ohta H, Sugimoto K, Masuda T. 2012. Regulation of root greening by light and auxin/cytokinin signaling in Arabidopsis. Plant Cell 24: 1081-1095.
- 37. Kobayashi K, Masuda T, Tajima N, Wada H, Sato N. 2014. Molecular phylogeny and intricate evolutionary history of the three isofunctional enzymes involved in the oxidation of protoporphyrinogen IX. Genome Biol Evol 6: 2141-2155.
- 38. Kobayashi K, Masuda T. 2016. Transcriptional regulation of tetrapyrrole biosynthesis in Arabidopsis thaliana. Front Plant Sci 7: 1811
- 39. Kobayashi , Koichi , Mochizuki N, Yoshimura N, Motohashi K, Hisabori T, Masuda T. 2008. Functional analysis of Arabidopsis thaliana isoforms of the Mg-chelatase CHLI subunit. Photochem Photobiol 7: 1188-1195.
- 40. Kong W, Yu X, Chen H, Liu L, Xiao Y, Wang Y, Wang C, Lin Y, Yu Y, Wang C, Jiang L, Zhai H, Zhao Z, Wan J. 2016. The catalytic subunit of magnesium-protoporphyrin IX monomethyl ester cyclase forms a chloroplast complex to regulate chlorophyll biosynthesis in rice. Plant Mol Biol 92: 177-191.
- 41. Kusaba M, Ito H, Morita R, Iida S, Sato Y, Fujimoto M, Kawasaki S, Tanaka R, Hirochika H, Nishimura M, Tanaka A. 2007. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell 19: 1362-1375.
- 42. Kusano M, Fukushima A, Tabuchi-Kobayashi M, Funayama K, Kojima S, Maruyama K, Yamamoto YY, Nishizawa T, Kobayashi M, Wakazaki M, Sato M, Toyooka K, Osanai-Kondo K, Utsumi Y, Seki M, Fukai C, Saito K, Yamaya T. 2020. Cytosolic GLUTAMINE SYNTHETASE1; 1 Modulates metabolism and chloroplast development in Roots. Plant Physiol 182: 1894-1909.
- 43. Lau OS, Deng XW. 2012. The photomorphogenic repressors COP1 and DET1: 20 years later. Trends Plant Sci 17: 584-593.
- 44. Lee J, He K, Stolc V, Lee H, Figueroa P, Gao Y, Tongprasit W, Zhao H, Lee I, Deng XW. 2007. Analysis of transcription factor HY5 genomic binding sites revealed its hierarchical role in light regulation of development. Plant Cell 19: 731-749.
- 45. Lee S, Kim JH, Yoo ES, Lee CH, Hirochika H, An G. 2005. Differential regulation of chlorophyll a oxygenase genes in rice. Plant Mol Biol 57: 805-818.
- 46. Legnaioli T, Cuevas J, Mas P. 2009. TOC1 functions as a molecular switch connecting the cir cadian clock with plant responses to drought. EMBO J 28: 3745-3757.
- 47. Leivar P, Quail PH. 2011. PIFs: pivotalc omponents in a cellular signaling hub. Trends Plant Sci 16: 19-28.
- 48. Leivar P, Tepperman JM, Monte E, Calderon RH, Liu TL, Quail PH. 2009. Definition of early transcriptional circuitry involved inlight-induced reversal of PIF-imposed repression of photomorphogenesis in young Arabidopsis seedlings. Plant Cell 21: 3535-3553.
- 49. Li P, Song A, Li Z, Fan F, Liang Y. 2017. Transcriptome analysis in leaves of rice (Oryza sativa) under high manganese stress. Biologia 724: 388-397.
- 50. Li RQ, Huang JZ, Zhao HJ, Fu HW, Li YF, Liu GZ, Shu QY. 2014. A down-regulated epi-allele of the genomes uncoupled 4 gene generates a xantha marker trait in rice. Theor Appl Genet 127: 2491-2501.
- 51. Li X, Duan X, Jiang H, Sun Y, Tang Y, Yuan Z, Guo J, Liang W, Chen L, Yin J, Ma H, Wang J, Zhang D. 2006. Genome-wide analysis of basic/helix-loop-helix transcription factor family in rice and Arabidopsis. Plant Physiol 141: 1167-1184.
- 52. Lim SH, Witty M, Wallace-Cook AD, Ilag LI, Smith AG. 1994. Porphobilinogen deaminase is encoded by a single gene in Arabidopsis thaliana and is targeted to the chloroplasts. Plant Mol Biol 26: 863-872.
- 53. Lin YP, Lee TY, Tanaka A, Charng YY. 2014. Analysis of an Arabidopsis heat-sensitive mutant reveals that chlorophyll synthase is involved in reutilization of chlorophyllide during chlorophyll turnover. Plant J 80: 14-26.
- 54. Liu X, Li Y, Zhong S. 2017. Interplay between light and plant hormones in the control of Arabidopsis seedling chlorophyll biosynthesis. Front Plant Sci 8: 1433
- 55. Ma Z, Hu X, Cai W, Huang W, Zhou X, Luo Q, Yang H, Wang J, Huang J. 2014. Arabidopsis miR171-Targeted Scarecrow-Like proteins bind to GT cis-elements and mediate gibberellin-regulated chlorophyll biosynthesis under light conditions. PLoS Genet 10: e1004519
- 56. Mach JM, Castillo AR, Hoogstraten R, Greenberg JT. 2001. The Arabidopsis-accelerated cell death gene ACD2 encodes red chlorophyll catabolite reductase and suppresses the spread of disease symptoms. Proc Natl Acad Sci USA 98: 771-776.
- 57. Mara CD, Irish VF. 2008. Two GATA transcription factors are downstream effectors of floral homeotic gene action in Arabidopsis. Plant Physiol 147: 707-718.
- 58. Matsumoto F, Obayashi T, Sasaki-sekimoto Y, Ohta H, Takamiya K. 2004. Gene expression profiling of the tetrapyrrole metabolic pathway in Arabidopsis with a mini-array system. Plant Physiol 135: 2379-2391.
- 59. McCormac AC, Fischer A, Kumar AM, Sö ll D, Terry MJ. 2001. Regulation of HEMA1 expression by phytochrome and a plastid signal during de-etiolation in Arabidopsis thaliana. Plant J 25: 549-561.
- 60. McCormac AC, Terry MJ. 2002. Light-signalling pathways leading to the co-ordinated expression of HEMA1 and Lhcb during chloroplast development in Arabidopsis thaliana. Plant J 32: 549-559.
- 61. Meguro M, Ito H, Takabayashi A, Tanaka R, Tanaka A. 2011. Identification of the 7-hydroxymethyl chlorophyll a reductase of the chlorophyll cycle in Arabidopsis. Plant Cell 23: 3442-3453.
- 62. Meskauskiene R, Klaus A. 2002. Interaction of FLU, a negative regulator of tetrapyrrole biosynthesis, with the glutamyl-tRNA reductase requires the tetratricopeptide repeat domain of FLU. FEBS Lett 532: 27-30.
- 63. Meskauskiene R, Nater M, Goslings D, Kessler F, op den Camp R, Apel K. 2001. FLU: a A negative regulator of chlorophyll biosynthesis in Arabidopsis thaliana. Proc Natl Acad Sci USA 98: 12826-12831.
- 64. Monte E, Tepperman JM, Al-Sady B, Kaczorowski KA, Alonso JM, Ecker JR, Li X, Zhang Y, Quail PH. 2004. The phytochrome-interacting transcription factor, PIF3, acts early, selectively, and positively in light-induced chloroplast development. Proc Natl Acad Sci USA 101: 16091-16098.
- 65. Moon J, Zhu L, Shen H, Huq E. 2008. PIF1 directly and indirectly regulates chlorophyll biosynthesis to optimize the greening process in Arabidopsis. Proc Natl Acad Sci USA 105: 9433-9438.
- 66. Morita R, Sato Y, Masuda Y, Nishimura M, Kusaba M. 2009. Defect in non-yellow coloring 3, an α/β hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice. Plant J 59: 940-952.
- 67. Murakami M, Tago Y, Yamashino T, Mizuno T. 2007. Characterization of the rice circadian clock-associated pseudo-response regulators in Arabidopsis thaliana. Biosci Biotechnol Biochem 71: 1107-1110.
- 68. Nagai S, Koide M, Takahashi S, Kikuta A, Aono M, Sasaki-Sekimoto Y, Ohta H, Takmiya K, Masuda T. 2007. Induction of isoforms of tetrapyrrole biosynthetic enzymes, AtHEMA2 and AtFC1, under stress conditions and their physiological functions in Arabidopsis. Plant Physiol 144: 1039-1051.
- 69. Nakamura H, Muramatsu M, Hakata M, Ueno O, Nagamura Y, Hirochika H, Takano M, Ichikawa H. 2009. Ectopic overexpression of the transcription factor OsGLK1 induces chloroplast development in non-green rice cells. Plant Cell Physiol 50: 1933-1949.
- 70. Nakamura Y, Kato T, Yamashino T, Murakami M, Mizuno T. 2007. Characterization of a set of phytochrome-interacting factor-like bHLH proteins in Oryza sativa. Biosci Biotechnol Biochem 71: 1183-1191.
- 71. Oberhuber M, Berghold J, Breuker K, Hörtensteiner S, Kraäüutler B. 2003. Breakdown of chlorophyll: A nonenzymatic reaction accounts for the formation of the cololess 'nonfluorescent' chlorophyll catabolites. Proc Natl Acad Sci USA 100: 6910-6915.
- 72. Oborník M, Green BR. 2005. Mosaic origin of the heme biosynthesis pathway in photosynthetic eukaryotes. Mol Biol Evol 22: 2343-2353.
- 73. op den Camp RGL, Przybyla D, Ochsenbein C, Laloi C, Kim C, Danon A, Wagner D, Hideg E, Göbel C, Feussner I, Nater M, Apel K. 2003. Rapid induction of distinct stress responses after the release of singlet oxygen in Arabidopsis. Plant Cell 15: 2320-2332.
- 74. Ouyang X, Li J, Li G, Li B, Chen B, Shen H, Huang X, Mo X, Wan X, Lin R, Li S, Wang H, Deng XW. 2011. Genome-wide binding site analysis of FAR-RED ELONGATED HYPOCOTYL 3 reveals its novel function in Arabidopsis development. Plant Cell 23: 2514-2535.
- 75. Papenbrock J, Mock H, Kruse E, Grimm B. 1999. Expression studies in tetrapyrrole biosynthesis: inverse maxima of magnesium chelatase and ferrochelatase activity during cyclic photoperiods. Planta 20849: 264-273.
- 76. Park MR, Baek SH, de Los Reyes BG, Yun SJ, Hasenstein KH. 2012. Transcriptome profiling characterizes phosphate deficiency effects on carbohydrate metabolism in rice leaves. J Plant Physiol 169: 193-205.
- 77. Parker R, Flowers TJ, Moore AL, Harpham NV. 2006. An accurate and reproducible method for proteome profiling of the effects of salt stress in the rice leaf lamina. J Exp Bot 57: 1109-1118.
- 78. Phung TH, Jung HI, Park JH, Kim JG, Back K, Jung S. 2011. Porphyrin biosynthesis control under water stress: sustained porphyrin status correlates with drought tolerance in transgenic rice. Plant physiol 157: 1746-1764.
- 79. Piao W, Han SH, Sakuraba Y, Paek NC. 2017. Rice 7-hydroxymethyl chlorophyll a reductase is involved in the promotion of chlorophyll degradation and modulates cell death signaling. Mol Cells 40: 773-786.
- 80. Piao W, Kim EY, Han SH, Sakuraba Y, Paek NC. 2015. Rice Phytochrome B (OsPhyB) negatively regulates dark- and starvation-induced leaf senescence. Plants (Basel) (Basel): 644-663.
- 81. Powell ALT, Nguyen CV, Hill T, Cheng KL, Figueroa-Balderas R, Aktas H, Ashrafi H, Pons C, Fernández-Muñoz R, Vicente A, Lopez-Baltazar J, Barry CS, Liu Y, Chetelat R, Granell A, Van Deynze A, Giovannoni JJ, Bennett AB. 2012. Uniform ripening encodes a Golden2-like transcription factor regulating tomato fruit chloroplast development. Science 336: 1711-1715.
- 82. Pruzinská A, Anders I, Aubry S, Schenk N, Tapernoux-Lüthi E, Müller T, Kräutler B, Hörtensteiner S. 2007. In vivo participation of red chlorophyll catabolite reductase in chlorophyll breakdown. Plant Cell 19: 369-387.
- 83. Pruzinská A, Tanner G, Anders I, Roca M, Hörtensteiner S. 2003. Chlorophyll breakdown: pheophorbide pheophorbide a oxygenase is a rieske-type iron-sulfur protein, encoded by the accelerated cell death 1 gene. Proc Natl Acad Sci USA 100: 15259-15264.
- 84. Quail PH. 2002. Photosensory perception and signalling in plant cells: new paradigms? Curr Opin Cell Biol 14: 180-188.
- 85. Reyes JC, Muro-Pastor MI, Florencio FJ. 2004. The GATA family of transcription factors in Arabidopsis and rice. Plant Physiol 134: 1718-1732.
- 86. Richter AS, Hochheuser C, Fufezan C, Heinze L, Kuhnert F, Grimm B. 2016. Phosphorylation of GENOMES UNCOUPLED 4 alters stimulation of Mg chelatase activity in angiosperms. Plant Physiol 172: 1578-1595.
- 87. Richter AS, Wang P, Grimm B. 2016. Arabidopsis Mg-Protoporphyrin IX Methyltransferase activity and redox regulation depend on conserved cysteines. Plant Cell Physiol 57: 519-527.
- 88. Richter R, Behringer C, Müller IK, Schwechheimer C. 2010. The GATA-type transcription factors GNC and GNL/CGA1 repress gibberellin signaling downstream from DELLA proteins and PHYTOCHROME-INTERACTING FACTORS. Genes Dev 24: 2093-2104.
- 89. Rodoni S, Mühlecker W, Anderl M, Kräutler B, Moser D, Thomas H, Matile P, Hörtensteiner S. 1997a. Chlorophyll breakdown in senescent chloroplasts (Cleavage of pheophorbide a in two enzymic steps). Plant Physiol 115: 669-676.
- 90. Rodoni S, Vicentini F, Schellenberg M, Matile P, Hörtensteiner S. 1997b. Partial purification and characterization of red chlorophyll catabolite reductase, a stroma protein involved in chlorophyll breakdown. Plant Physiol 115: 677-682.
- 91. Rong H, Tang Y, Zhang H, Wu P, Chen Y, Li M, Wu G, Jiang H. 2013. The Stay-Green Rice like (SGRL) gene regulates chlorophyll degradation in rice. Plant Physiol 170: 1367-1373.
- 92. Rzeznicka K, Walker CJ, Westergren T, Kannangara CG, von Wettstein D, Merchant S, Gough SP, Hansson M. 2005. Xantha-l encodes a membrane subunit of the aerobic Mg-protoporphyrin IX monomethyl ester cyclase involved in chlorophyll biosynthesis. Proc Natl Acad Sci USA 102: 5886-5891.
- 93. Sakuraba Y, Park SY, Kim YS, Wang SH, Yoo SC, Hörtensteiner S, Paek NC. 2014. Arabidopsis STAY-GREEN2 is a negative regulator of chlorophyll degradation during leaf senescence. Mol Plant 7: 1288-1302.
- 94. Sakuraba Y, Rahman ML, Cho SH, Kim YS, Koh HJ, Yoo SC, Paek NC. 2013. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J 74: 122-133.
- 95. Sato Y, Morita R, Katsuma S, Nishimura M, Tanaka A, Kusaba M. 2009. Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J 57: 120-131.
- 96. Sato Y, Morita R, Nishimura M, Yamaguchi H, Kusaba M. 2007. Mendel's green cotyledon gene encodes a positive regulator of the chlorophyll-degrading pathway. Proc Natl Acad Sci USA 104: 14169-14174.
- 97. Scharfenberg M, Mittermayr L, von Roepenack-Lahaye E, Schlicke H, Grimm B, Leister D, Kleine T. 2015. Functional characterization of the two ferrochelatases in Arabidopsis thaliana. Plant Cell Environ 38: 280-298.
- 98. Schelbert S, Aubry S, Burla B, Agne B, Kessler F, Krupinska K, Hörtensteiner S. 2009. Pheophytin pheophorbide hydrolase (pheophytinase) is involved in chlorophyll breakdown during leaf senescence in Arabidopsis. The Plant Cell 21: 767-785.
- 99. Scheumann V, Ito H, Tanaka A, Schoch S, Rüdiger W. 1996. Substrate specificity of chlorophyll (ide) b reductase in etioplasts of barley (Hordeum vulgare L.). Eur J Biochem 242: 163-170.
- 100. Scheumann V, Schoch S, Rüdiger W. 1998. Chlorophyll a formation in the chlorophyll b reductase reaction requires reduced ferredoxin. J Biol Chem 273: 35102-35108.
- 101. Scheumann V, Schoch S, Rüdiger W. 1999. Chlorophyll b reduction during senescence of barley seedlings. Planta 209: 364-370.
- 102. Schlicke H, Salinas AHartwig AS, Firtzlaff V, Richter AS, Glässer C, Maier K, Finkemeier I, Grimm B. 2014. Induced deactivation of genes encoding chlorophyll biosynthesis enzymes disentangles tetrapyrrole-mediated retrograde signaling. Mol Plant 7: 1211-1227.
- 103. Shalygo N, Czarnecki O, Peter E, Grimm B. 2009. Expression of chlorophyll synthase is also involved in feedback-control of chlorophyll biosynthesis. Plant Mol Biol 71: 425-436.
- 104. Shimoda Y, Ito H, Tanaka A. 2016. Arabidopsis STAY-GREEN, mendel's green cotyledon gene, encodes magnesium-dechelatase. Plant Cell 28: 2147-2160.
- 105. Shin J, Kim K, Kang H, Zulfugarov IS, Bae G, Lee CH, Lee D, Choi G. 2009. Phytochromes promote seedling light responses by inhibiting four negatively acting phytochrome-interacting factors. Proc Natl Acad Sci USA 106: 7660-7665.
- 106. Stephenson PG, Fankhauser C, Terry MJ. 2009. PIF3 is a repressor of chloroplast development. Proc Natl Acad Sci USA 106: 7654-7659.
- 107. Stephenson PG, Terry MJ. 2008. Light signalling pathways regulating the Mg-chelatase branch point of chlorophyll synthesis during de-etiolation in Arabidopsis thaliana. Photochem Photobiol Sci 7: 1243-1252.
- 108. Sun C, Liu L, Tang J, Lin A, Zhang F, Fang J, Zhang G, Chu C. 2011. RLIN1, encoding a putative coproporphyrinogen III oxidase, is involved in lesion initiation in rice. J Genet Genomics 38: 29-37.
- 109. Takahashi K, Takabayashi A, Tanaka A, Tanaka R. 2014. Functional analysis of light-harvesting-like protein 3 (LIL3) and its light-harvesting chlorophyll-binding motif in Arabidopsis. J Biol Chem 2892: 987-999.
- 110. Tamai H, Iwabuchi M, Meshi T. 2002. Arabidopsis GARP transcriptional activators interact with the Pro-rich activation domain shared by G-box-binding bZIP factors. Plant Cell Physiol 43: 99-107.
- 111. Tan FC, Cheng Q, Saha K, Heinemann IU, Jahn M, Jahn D, Smith AG. 2008. Identification and characterization of the Arabidopsis gene encoding the tetrapyrrole biosynthesis enzyme uroporphyrinogen III synthase. Biochem J 410: 291-299.
- 112. Tanaka A, Ito H, Tanaka R, Tanaka NK, Yoshida K, Okada K. 1998. Chlorophyll a oxygenase (CAO) is involved in chlorophyll b formation from chlorophyll a. Proc Natl Acad Sci USA 95: 12719-12723.
- 113. Tanaka R, Kobayashi K, Masuda T. 2011. Tetrapyrrole metabolism in Arabidopsis thaliana. Arabidopsis Book 9: e0145
- 114. Tang W, Wang W, Chen D, Ji Q, Jing Y, Wang H, Lin R. 2012. Transposase-derived proteins FHY3/FAR1 interact with PHYTOCHROME-INTERACTING FACTOR 1 to regulate chlorophyll biosynthesis by modulating HEMB1 during deetiolationin Arabidopsis. Plant Cell 24: 1984-2000.
- 115. Tang Y, Li M, Chen Y, Wu P, Wu G, Jiang H. 2011. Knockdown of OsPAO and OsRCCR1 cause different plant death phenotypes in rice. Plant Physiol 168: 1952-1959.
- 116. Toledo-Ortiz G, Huq E, Rodríiguez-Concepcióon M. 2010. Direct regulation of phytoene synthase gene expression and carotenoid biosynthesis by phytochrome-interacting factors. Proc Natl Acad Sci USA 107: 11626-11631.
- 117. Toledo-Ortiz G, Johansson H, Lee KP, Bou-Torrent J, Stewart K, Steel G, Rodríguez-Concepción M, Halliday KJ. 2014. The HY5-PIF regulatory module coordinates light and temperature control of photosynthetic gene transcription. PLoS Genet 10: e1004416
- 118. Triantaphylidès C, Havaux M. 2009. Singlet oxygen in plants: production, detoxification and signaling. Trends Plant Sci 14: 219-228.
- 119. Tsuchiya T, Ohta H, Okawa K, Iwamatsu A, Shimada H, Masuda T, Takamiya K. 1999. Cloning of chlorophyllase, the key enzyme in chlorophyll degradation: finding of a lipase motif and the induction by methyl jasmonate. Proc Natl Acad Sci USA 96: 15362-15367.
- 120. Tsuchiya T, Suzuki T, Yamada T, Shimada H, Masuda T, Ohta H, Takamiya K. 2003. Chlorophyllase as a serine hydrolase: identification of a putative catalytic triad. Plant Cell Physiol 44: 96-101.
- 121. von Arnim A, Deng XW. 1996. Light control of seedling development. Annu Rev Plant Physiol Plant Mol Biol 47: 215-243.
- 122. Wang H, Wang H. 2015. Multifaceted roles of FHY3 and FAR1 in light signaling and beyond. Trends Plant Sci 20: 453-461.
- 123. Wang L, Mai YX, Zhang YC, Luo Q, Yang HQ. 2010. MicroRNA 171c-targeted SCL6-II, SCL6-III, and SCL6-IV genes regulate shoot branching in Arabidopsis. Mol Plant 3: 794-806.
- 124. Wang P, Fouracre J, Kelly S, Karki S, Gowik U, Aubry S, Shaw MK, Peter W, Slamet-Loedin IH, Quick WP, Hibberd JM, Langdale JA. 2013. Evolution of GOLDEN2-LIKE gene function in C3 and C4 plants. Planta 237: 481-495.
- 125. Wang P, Gao J, Wan C, Zhang F, Xu Z, Huang X, Sun X, Deng X. 2010. Divinyl chlorophyll (ide) a can be converted to monovinyl chlorophyll (ide) a by a divinyl reductase in rice. Plant Physiol 153: 994-1003.
- 126. Wang X, Huang R, Quan R. 2017. Mutation in Mg-protoporphyrin IX monomethyl ester cyclase decreases photosynthesis capacity in rice. PloS One 12: e0171118
- 127. Wang Z, Hong X, Hu K, Wang Y, Wang X, Du S, Li Y, Hu D, Chen K, An B, Li Y. 2017. Impaired magnesium protoporphyrin IX methyltransferase (ChlM) impedes chlorophyll synthesis and plant growth in rice. Front Plant Sci 8: 1694
- 128. Waters MT, Wang P, Korkaric M, Capper RG, Saunders NJ, Langdale JA. 2009. GLK transcription factors coordinate expression of the photosynthetic apparatus in Arabidopsis. Plant Cell 21: 1109-1128.
- 129. Woodson JD, Joens MS, Sinson AB, Gilkerson J, Salomé PA, Weigel D, Fitzpatrick JA, Chory J. 2015. Ubiquitin facilitates a quality-control pathway that removes damaged chloroplasts. Science 350: 450-454.
- 130. Wüthrich KL, Bovet L, Hunziker PE, Donnison IS, Hörtensteiner S. 2000. Molecular cloning, functional expression and characterisation of RCC reductase involved in chlorophyll catabolism. Plant J 21: 189-198.
- 131. Xu G, Guo H, Zhang D, Chen D, Jiang Z, Lin R. 2015. REVEILLE1 promotes NADPH: protochlorophyllide oxidoreductase A expression and seedling greening in Arabidopsis. Photosynth Res 126: 331-340.
- 132. Zeng Y, Ma L, Ji Z, Wen Z, Li X, Shi C, Yang C. 2013. Fine mapping and candidate gene analysis of LM3, a novel lesion mimic gene in rice. Biologia 68: 82-90.
- 133. Zeng ZQ, Lin TZ, Zhao JY, Zheng TH, Xu LF, Wang YH, Liu LL, Jiang L, Chen SH, Wan JM. 2020. OsHemA gene, encoding glutamyl-tRNA reductase (GluTR) is essential for chlorophyll biosynthesis in rice (Oryza sativa). J Integr Agri 19: 612-623.
- 134. Zhang F, Tang W, Hedtke B, Zhong L, Liu L, Peng L, Lu C, Grimm B, Lin R. 2014. Tetrapyrrole biosynthetic enzyme protoporphyrinogen IX oxidase 1 is required for plastid RNA editing. Proc Natl Acad Sci USA 111: 2023-2028.
- 135. Zhang H, Li J, Yoo JH, Yoo SC, Cho SH, Koh HJ, Seo HS, Paek NC. 2006. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol 62: 325-337.
- 136. Zhao Y, Han Q, Ding C, Huang Y, Liao J, Chen T, Feng S, Zhou L, Zhang Z, Chen Y, Yuan S, Yuan M. 2020. Effect of low temperature on chlorophyll biosynthesis and chloroplast biogenesis of rice seedlings during greening. Int J Mol Sci 21: 1390
- 137. Zhou F, Wang CY, Gutensohn M, Jiang L, Zhang P, Zhang D, Dudareva N, Lu S. 2017. A recruiting protein of geranylgeranyl diphosphate synthase controls metabolic flux toward chlorophyll biosynthesis in rice. Proc Natl Acad Sci USA 114: 6866-6871.
- 138. Zhou J, Liu Q, Zhang F, Wang Y, Zhang S, Cheng H, Yan L, Li L, Chen F, Xie X. 2014. Overexpression of OsPIL15, a phytochrome-interacting factor-like protein gene, represses etiolated seedling growth in rice. J Integr Plant Biol 56: 373-387.
- 139. Zhu L, Bu Q, Xu X, Paik I, Huang X, Hoecker U, Deng XW, Huq E. 2015. CUL4 forms an E3 ligase with COP1 and SPA to promote light-induced degradation of PIF1. Nat Commun 6: 1-10.