적요
Enhancing pod shatter resistance is essential for improving the mechanical harvesting efficiency of rapeseed (Brassica napus L.). This study aimed to evaluate pod shatter resistance in domestic breeding lines and genetic resources, as well as to investigate the relationship between shatter resistance and agronomic traits thereby providing foundational data for the development of shatter-resistant cultivars. Pod shatter resistance was assessed using the Random Impact Test for 105 accessions in 2022–2023 and 159 accessions in 2023–2024, with the Silique Shatter Resistance Index (SSRI) as the evaluation criterion. Most accessions exhibited susceptibility to pod shatter, whereas IT031375 showed high shatter resistance, identifying it as a promising resource for the development of shatter-resistant cultivars. Analysis of the relationship between weather conditions and pod shatter resistance revealed that longer sunlight exposure and lower precipitation during the growing stages contributed to improved pod development and shatter resistance. Furthermore, a correlation analysis between SSRI and agronomic traits demonstrated significant positive correlations with silique length (r=0.368, p<0.001) and weight (r=0.451, p<0.001). Increased silique length and weight are associated with enhanced pod wall development and seed weight, which are critical for improving yield and shatter resistance.
서언
유채(
Brassica napus L., AACC)는 기름, 식품, 경관 조성 등 다양한 용도로 활용되는 중요한 작물이다(
Nath et al. 2016). 유채유는 포화지방산 함량이 낮고 불포화지방산이 풍부하며, 토코페롤 및 피토스테롤을 함유하여 건강에 유익한 식용유로 알려져 있으며, 전 세계에서 세 번째로 많이 생산되는 식용 유지이다(
Piao et al. 2013,
Shen et al. 2023). 국내에서는 유채를 주로 경관 용도로 재배하고 꽃이 핀 직후 제거하였으나, 최근에는 겨울철 유휴 농경지의 활용과 유채유의 우수성에 대한 인식증가로 종자를 수확하여 유채유를 생산하는 재배단지가 확대되고 있다(
Jeong et al. 2024).
유채의 기계화 재배는 노동력을 절감하고 생산성을 높이는 중요한 기술이지만, 기계 수확 과정에서 발생하는 수량 손실은 해결해야 할 과제이다. 유채 종자 손실의 주요 원인은 수확 전이나 수확 중에 꼬투리가 충격을 받아 종자가 배출되는 꼬투리 탈립(Silique shatter)이다. 유채 꼬투리의 탈립은 리그닌화된 세포와 비리그닌화된 세포 간의 수축 차이로 인한 내부 장력 발생과 다당류 분해 효소(Polygalacturonase, Cellulase)의 작용으로 세포가 분리되며 발생하며(
Meakin & Roberts 1990a), 꼬투리 성숙 후기에는 중축 세포의 균열로 인해 영양물질 수송이 차단되면서 분리가 가속화된다(
Meakin & Roberts 1990b). 탈립은 꼬투리가 성숙되었을 때 강풍이나 강우로 인한 꼬투리와 꼬투리간, 꼬투리와 줄기의 충돌로 발생하며, 수확 중에는 수확 기계와 꼬투리 간의 충돌로 인한 물리적 손상으로 발생한다. 이러한 꼬투리 탈립으로 인해 발생하는 종자 손실은 전체 수확량의 약 10%를 차지하며, 기상 조건이 불리할 경우 20% 이상 발생할 수 있다(
Price et al. 1996). 수량 손실 뿐만 아니라, 지면에 떨어진 유채 종자는 자생 유채(Volunteer rapeseed)로 자라 후작물에 잡초로서 부정적인 영향을 미칠 수 있고, 다른 유채 품종과의 교잡을 통해 유전적 오염을 일으킬 가능성도 있다(
Morgan et al. 1998). 꼬투리 탈립에 의해 종자 손실이 일어나는 주요 원인은 유채의 성숙이 일관되지 않기 때문이다. 유채 꼬투리 성숙기는 개화 후 약 60일이며, 적정 기계 수확 시기는 3~4일로 매우 짧다. 수확 시기가 늦어질 경우 과잉 성숙으로 물리적 충격에 약화되어 종자 손실이 증가하며, 적정 수확 시기보다 너무 일찍 수확하면 줄기와 꼬투리의 수분함량이 높아 수확 기계의 성능과 수확 효율이 저하된다. 또한, 미성숙한 종자로부터 추출된 기름은 클로로필에 오염되어 품질 저하를 초래한다(
Hu et al. 2015). 따라서 적정 수확 시기의 연장을 위해 탈립 저항성을 높이는 것은 수량 손실 감소와 품질 향상에 기여할 수 있다.
탈립 저항성을 강화한 품종은 적정 수확 시기를 연장하여 성숙 균일도를 높이고 탈립에 의한 종자 손실을 줄일 뿐만 아니라, 기름 함량도 증가시켜 경제적 효율성을 높일 수 있다. 그러나 국내에서는 유채 내탈립성 품종 육성에 대한 연구가 부족한 실정이다. 따라서 국내 육성 품종, 계통 및 유전자원을 활용한 탈립 저항성 스크리닝과 꼬투리 특성 간 관계 구명 연구가 필요하다. 본 연구는 2022-2023년과 2023-2024년 동안 내탈립성 품종 육성을 위해 탈립 저항성을 평가하여 내탈립 자원을 선발하고, 기상 조건에 따른 변화를 분석하였다. 또한, 꼬투리 특성과 탈립 저항성 간의 상관관계를 분석하여 내탈립성 계통 선발을 위한 지표를 설정하고자 하였다.
재료 및 방법
식물 재료 및 재배 조건
본 연구는 국립식량과학원 바이오에너지작물연구소에서 육성한 유채 품종과 계통, 그리고 유전자원센터에서 분양받은 자원을 대상으로 하였다. 시험은 2022-2023년 및 2023-2024년에 바이오에너지작물연구소에서 수행되었으며, 각각 105자원(2023년)과 159자원(2024년)을 평가하였고, 이 중 28자원은 2년 동안 반복 시험을 실시하였다(
Supplementary Table 1). 시험구는 구획당 면적 4.0 m
2 (2 m×2 m)이며, 5줄로 구성되었고, 식물 간격은 40 cm로 설정하였다. 2023년에는 9월 22일, 2024년에는 9월 27일에 육묘판에 파종을 하였으며 본엽 4매 상태가 되었을 때 이식하였다. 비료 시비량은 10 a당 N-P
2O
5-K
20를 기준으로 기비 7.6-40.0-13.3와 퇴비 1,000 kg를 시용하였다. 추비는 요소(46% N)를 14.1 kg를 시용하였다. 기비와 퇴비는 이식하기 2일 전, 추비는 생육재생기 이후에 시용하였다. 병해충 관리 및 생육 관리는 농촌진흥청 유채 표준재배법에 따라 수행하였다(
RDA 2012). 생육 단계별 기상 온도, 강수량, 일조시간은 농촌진흥청 농업기상관측시스템을 이용해 측정하였다(
Table 1).
개화 후 60일이 지나 생리적 성숙기에 도달한 꼬투리를 각 자원별로 주경화서(Main inflorescence)에서 채취한 후 자연 건조(Air-dried)한 뒤 탈립 저항성을 측정하였다. 탈립 저항성은 꼬투리와 외부요인과의 충돌하는 상황을 모방하여, 쇠구슬과 유채 꼬투리에 진동을 주어 탈립 저항성을 측정하기 위하여 주로 사용되는 Random impact test (RIT)를 기반으로 하여 수정된 방법을 이용해 측정하였다(
Bruce et al. 2002,
Hu et al. 2012,
Hu et al. 2015,
Liu et al. 2016,
Morgan et al. 1998). 내부 지름 20 cm, 높이 12 cm의 플라스틱 용기에 직경 8 mm의 쇠구슬 50개를 넣고, 다단 진탕 배양기(Visonbionex, VS-8480MX2-DT, Bucheon, Republic of Korea)를 사용하여 280 rpm으로 2분간 총 5회 진탕하였다. 각 진탕 횟수마다 열개된 꼬투리 수를 측정하였으며, 2반복으로 진행하였다. 열개된 꼬투리는 꽃자루(Pedicel)나 꼬투리 끝 부위(Beak)의 꼬투리 벽(Silique wall)이 분리되어 종자가 배출된 경우를 기준으로 하였다. 꼬투리 탈립 저항성 지수(Silique shatter resistance index, SSRI)는 다음 식으로 계산하였다:
xi: i번째(1≤i≤5) 진탕에서 열개된 꼬투리 수
n1: 총 꼬투리 수(20개)
n2: 진탕 회수(5회)
꼬투리 농업적 특성 조사
꼬투리의 농업적 특성은 20개의 꼬투리를 이용하여 측정하였다. 꼬투리 길이(Silique length)는 꽃자루 길이(Pedicel length), 꼬투리 끝 부위 길이(Beak length)를 제외한 길이로 측정하였으며, 각각의 길이는 따로 측정하였다. 꼬투리 두께(Silique thickness), 꼬투리 벽 두께(Silique wall thickness), 격막 두께(Septum thickness)와 꽃자루-격막 접합부 두께(Pedicel-septum juction thickness)는 꼬투리 중간 부분에서 측정하였다. 또한, 20개 꼬투리의 총 무게(20-silique weight), 종자 무게(20-silique seed weight), 벽 무게(20-silique wall weight)도 측정하였다. 그 후, 꼬투리 특성과 탈립 저항성 지수 간의 피어슨 상관관계를 분석하였다.
통계 분석
모든 통계 분석은 Rstudio (verson 4.4.1; Rstudio, PBC, Boston, MA, USA)를 사용하였으며, LSD 검정과 상관관계 분석은 agricolae, GGally, ggplot2, dplyr 패캐지를 활용하여 수행하였다.
결과 및 고찰
유채 자원별 꼬투리 탈립 저항성 측정
2023년과 2024년에 Random Impact Test (RIT)를 이용하여 유채 자원의 꼬투리 탈립 저항성 지수(Silique shatter resistance index, SSRI)를 측정하였다(
Fig. 1). 2023의 SSRI는 0.005-0.895 범위로 나타났으며, 탈립 취약성 자원(Susceptible to shattering, SSRI 0.0-0.3)이 전체의 52.4%, 중간 취약성 자원(Moderate susceptible to shattering, SSRI 0.3-0.7)이 40.0%, 탈립 저항성 자원(Resistance to shattering, SSRI 0.7-1.0)이 7.6%로 분포하였다. 2024년의 SSRI는 0.01-0.845 범위를 보였으며, 탈립 취약성 자원이 70.4%, 중간 취약성 자원이 29.0%, 탈립 저항성 자원이 0.6%로 분포하였다.
두 해 모두 SSRI는 유사한 경향을 보였으며, 대부분의 자원이 탈립에 취약한 특성을 나타냈다. 특히, 국내 육성 품종의 SSRI는 모두 탈립 취약성 범위에 속해, 탈립 저항성 개선이 필요한 주요 특성으로 평가되었다(
Table 2). 반면, IT031375은 상대적으로 높은 탈립 저항성을 보여, 내탈립 품종 육성을 위한 유망 자원으로 확인되었다.
2022-2023년과 2023-2024년 동안의 기상 환경을 유채 생육 단계별로 분석하였다(
Table 1). 전체 생육 기간 동안 평균 기온과 총 강수량은 2023-2024년이 더 높았고, 총 일조시간은 2022-2023년이 더 길었다. 생육 단계별로 살펴보면, 2023-2024년은 2022-2023년보다 파종기부터 월동기, 월동기부터 착협기까지 강수량이 많고 일조시간이 짧았던 반면, 착협기부터 성숙기까지는 반대의 경향을 나타냈다.
동일한 28자원에 대해 SSRI를 측정한 결과, 2023년의 평균 SSRI (0.37)가 2024년(0.26)보다 유의미하게 높았다(
Table 2). 이러한 결과는 기상 환경이 유채 꼬투리의 발달 및 탈립 저항성에 중요한 영향을 미친다는 것을 보여준다.
Weymann et al. (2015)은 꼬투리 및 종자 형성기 동안의 기상 조건이 동화산물 가용성에 영향을 미쳐 종자의 형성과 발달에 중요한 역할을 한다고 보고하였다. 또한
Kuai et al. (2016)은 유묘기와 꼬투리 발달기에서 긴 일조시간이 탄수화물 축적을 촉진하여 꼬투리 발달을 강화한다고 하였다. 따라서, 2022-2023년의 종자 및 꼬투리 형성 기간 동안 긴 일조시간과 적은 강수량으로 2023-2024년에 비해 꼬투리 발달과 탈립 저항성 향상에 유리한 환경이 조성된 것으로 판단된다.
이와 같은 결과는 유채 생육 단계별 기상환경이 꼬투리 발달과 탈립 저항성에 상호적인 영향을 미친다는 것을 시사한다. 특히, 생육 초기와 꼬투리 형성기 동안의 일조시간과 강수량은 꼬투리 구조 형성과 탈립 저항성에 중요한 요인으로 작용한다. 따라서 꼬투리 탈립 저항성을 개선하기 위해서는 기상 환경과 생육 단계별 영향을 고려한 재배 관리가 필요할 것으로 사료된다.
꼬투리 탈립 저항성과 꼬투리 농업적 특성간의 상관분석
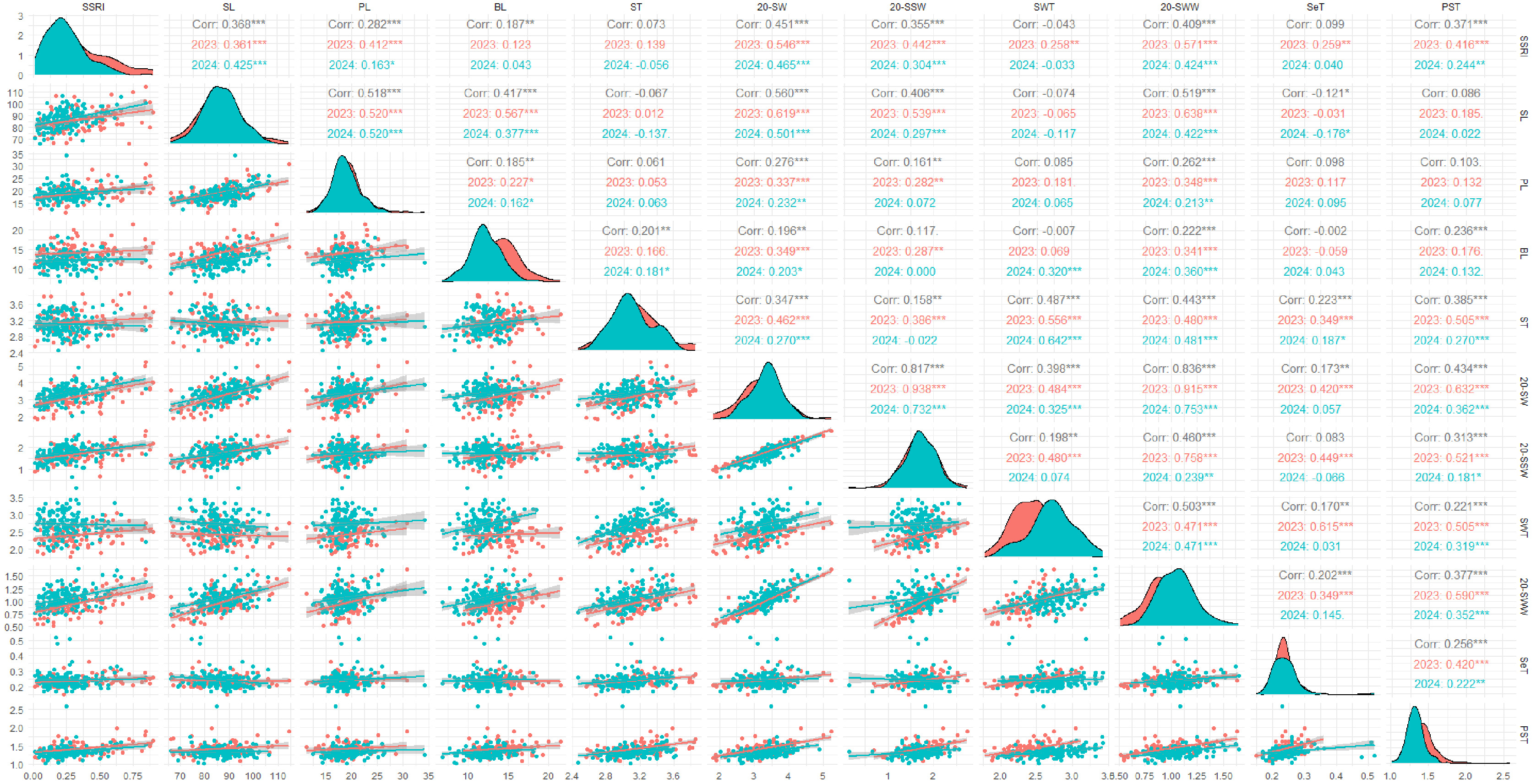
2023년과 2024년, 유채 꼬투리의 농업적 특성과 탈립 저항성(SSRI) 간의 관계를 분석하였다(
Table 3,
Fig. 2). 꼬투리 두께, 벽 두께, 격막 두께는 SSRI와 유의미한 상관관계를 보이지 않았다. 그러나, 꼬투리 길이와 SSRI는 양의 상관관계(r=0.368,
p<0.001)를 나타내었으며, 이는
Braatz et al. (2018),
Liu et al. (2013),
Qing et al. (2021)의 결과와 일치하고,
Wang et al. (2007) 및
Morgan et al. (1998)의 결과와는 반대였다. 꽃자루 길이와 꽃자루-격막 접합 두께 또한 SSRI와 유의미한 양의 상관관계를 보였다.
Hu et al. (2015)는 꽃자루(Pedicel) 부근의 격막축(Replum)이 두꺼울수록 탈립 저항성이 높아지며, 이는 탈리대(Dehiscence zone, DZ) 접합 부위 면적 증가와 관련이 있다고 보고하였다. 또한 꼬투리 길이가 길수록 격막축과 밸브 접합 부위가 증가하여 탈립 저항성이 향상된다고 하였다. 이러한 결과를 종합하면, 꼬투리 길이, 꽃자루 길이, 꽃자루-격막 접합부 두께의 증가는 관다발 조직 발달을 통해 꼬투리 탈립 저항성을 높이는 주요 요인임을 시사한다.
Qing et al. (2021)과
Wang et al. (2007)은 꼬투리 끝 부위 길이가 탈립 저항성과 관련이 없다고 보고하였는데, 본 연구에서는 연도별 꼬투리 끝 부위와 SSRI 간 상관관계가 각각 2023년 0.123 (
p>0.05), 2024년 0.043 (
p>0.05)로 유의미하지 않았다. 그러나 두 연도의 데이터를 통합하여 분석한 결과, 상관계수는 0.187 (
p<0.01)로 유의미한 수준을 보였으나, 이는 데이터 통합으로 샘플 크기가 증가하고 연도별 데이터 분포 차이가 반영된 결과로 사료되며, 추가 연구가 필요할 것으로 판단되었다.
20개 꼬투리 무게, 20개 종자 무게, 20개 벽 무게와 SSRI 간 상관계수는 각각 0.451, 0.355, 0.409 (
p<0.001)로 모두 유의미한 양의 상관관계를 나타냈다. 이는
Morgan et al. (1998),
Qing et al. (2021)의 결과와 일치하며, 꼬투리 벽 무게의 증가는 꼬투리의 생장, 발달 및 구조적 안정성을 높여 탈립 저항성을 강화한다고 보고된 바 있다(
Kuai et al. 2016). 20개 꼬투리의 무게를 구성하는 요소인 종자 무게와 벽 무게는 각각 r=0.817 및 r=0.836 (
p<0.001)의 높은 상관관계를 나타내었으며, 세 특성은 서로 밀접한 연관성을 보였다. 이러한 연관성은 꼬투리 무게, 종자 무게, 벽 무게의 증대가 꼬투리 탈립 저항성 증가에 기여할 수 있음을 나타낸다.
또한, 꼬투리 길이는 20개 꼬투리 무게, 종자 무게, 벽 무게와 양의 상관관계 (p<0.001)를 보여, 꼬투리 길이가 증가하면 종자 수와 벽 길이가 증가하여 무게가 증가하는 것을 확인하였다. 이러한 특성은 꼬투리 발달 과정에서 나타나는 결과로, 꼬투리 길이 증가가 내탈립성과 수확량에 모두 기여할 수 있음을 시사한다.
28자원에 대한 SSRI를 측정한 결과, 그룹 A (16자원)는 2023년의 SSRI가 2024년보다 유의미하게 낮아진 반면, 그룹 B는 반대의 경향을 나타냈다(
Table 2). 이러한 결과는 기상 환경과 더불어, SSRI와 양의 상관관계를 가지는 주요 농업적 형질이 연도별로 어떻게 변화하는지에 대한 분석이 필요하다는 것을 나타낸다. 이에 따라, SSRI와 상관관계를 가지는 형질들을 중심으로 그룹별 꼬투리 농업적 특성을 비교 분석하였다(
Table 4). 그룹 A의 경우, 2023년의 SSRI 값이 2024년보다 통계적으로 유의하게 높았다. SSRI와 상관성이 높은 형질들을 비교한 결과, 모든 주요 형질들이 2023년에서 전반적으로 더 높은 값을 보였다. 그러나 20개 종자 무게를 제외한 모든 형질에서 통계적 유의성은 나타나지 않았다. 반면, 그룹 B에서는 2023년보다 2024년에 SSRI 값이 증가하는 경향을 보였다. 꼬투리 길이, 20개 꼬투리 무게, 20개 꼬투리 벽 무게에서 2024년에 증가하는 양상을 나타냈으며, 특히 20개 꼬투리 벽 무게에서 통계적으로 유의한 증가가 확인되었다. 결론적으로, SSRI 값이 높게 나타난 연도에서는 SSRI와 양의 상관관계를 가지는 농업적 형질의 값 또한 상대적으로 높게 나타나는 경향을 보였다. 그러나, 본 연구에서 분석된 자원의 수는 제한적이었으며, 그룹 A와 그룹 B내 개별 형질 간 차이가 통계적으로 유의하지 않은 경우도 있었다. 따라서, 향후 더 많은 자원을 활용한 추가적인 연구가 필요하며, 기상조건과 농업적 형질 간의 상호작용에 대한 분석이 요구된다.
본 연구는 유채 꼬투리의 길이, 꽃자루 길이, 꽃자루-격막 접합부 두께, 그리고 꼬투리 무게(종자 및 벽 무게 포함)가 탈립 저항성과 유의미한 상관관계를 가짐을 확인하였다. 최근 연구들은 꼬투리 길이와 무게가 탈립 저항성을 강화한다고 보고하고 있으며(
Braatz et al. 2018,
Kuai et al. 2016,
Qing et al. 2021), 이러한 특성들은 내탈립 품종 선발의 신뢰할 수 있는 지표로 활용될 수 있다. 특히, 꼬투리 길이는 다른 주요 특성과도 밀접한 양의 상관관계를 보이며, 향후에는 꼬투리 발달 과정에서의 생리적 및 유전적 메커니즘을 구명하는 연구가 필요하다.
적요
유채의 기계 수확 효율성을 높이기 위해 꼬투리 탈립 저항성 강화가 필수적이다. 본 연구는 국내 육성 품종과 유전자원을 대상으로 꼬투리 탈립 저항성을 평가하고, 탈립 저항성과 농업적 특성 간의 관계를 구명하여 내탈립 품종 육성을 위한 기초자료를 제공하고자 수행되었다. 2022-2023년과 2023-2024년에 각각 105자원과 159자원을 대상으로 Random Impact Test (RIT)를 이용해 꼬투리 탈립 저항성 지수(Silique Shatter Resistance Index, SSRI)를 측정하였다. 대부분의 자원들은 탈립에 취약한 특성을 나타냈으며, IT031375은 높은 탈립 저항성을 가져 내탈립 품종 육성을 위한 유망 자원으로 확인되었다. 기상 환경과 꼬투리 탈립 저항성 간의 관계를 분석한 결과, 생육 단계에서 더 긴 일조시간과 낮은 강수량이 꼬투리 발달과 탈립 저항성 향상에 기여하는 것으로 나타났다. 또한, 꼬투리 농업적 특성과 SSRI 간의 상관분석 결과, 꼬투리 길이(r=0.368, p<0.001), 꼬투리 무게(r=0.451, p<0.001)와 SSRI 간에 유의미한 양의 상관관계를 나타냈다. 특히, 꼬투리 길이와 무게 증가는 꼬투리 벽 발달과 종자 무게 증가를 통해 수확량 증대와 탈립 저항성 향성에 중요한 역할을 하는 것으로 확인되었다.
보충자료
본문의 Supplementary Table 1은 한국육종학회지 홈페이지에서 확인할 수 있습니다.
사사
본 연구는 농촌진흥청 국립식량과학원 연구사업(과제명:유채 기계수확 적응 재배법 개선 연구(2세부), 과제번호: PJ017228022024)의 지원으로 수행되었습니다.
Fig. 1Phenotypic distribution for individual silique shatter resistance index (SSRI) in resources across 2 year (2023 and 2024). Y axis represents number of resources and×axis represents silique shatter resistance index (SSRI) measured by random impact test (RIT) method.

Fig. 2Correlation coefficients between silique shatter resistance index (SSRI) and silique agronomic traits. 2023: 105 genetic resources, 2024: 159 genetic resources. SSRI=silique shatter resistance index, SL=silique length, PL=pedicel length, BL=beack length, ST=silique thickness, 20-SW=20-silique weight, 20-SSW=20-silique seed weight, SWT=silique wall thickness, 20-SWW=20-silique wall wegiht, SeT=Septum thickness, PST=pedicel-septum juction thickness.
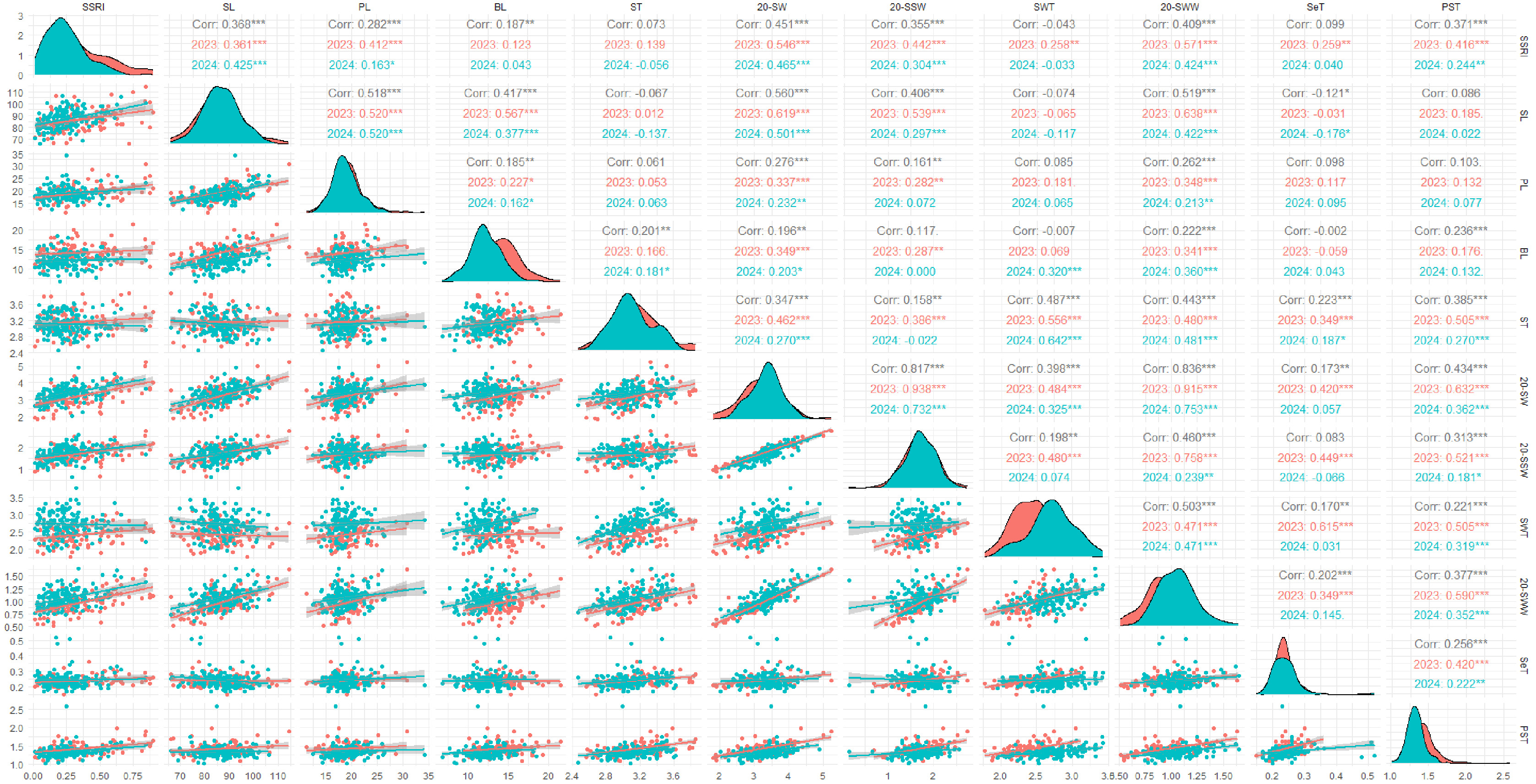
Table 1Meteorological conditions during the rapeseed growing seasons in 2022-2024.
Table 1
|
Growth stages |
Climatic index |
2022-2023 |
2023-2024 |
|
Seeding to wintering |
Average temperature (℃) |
6.4 |
7.2 |
|
Precipitation (mm) |
98.0 |
148.5 |
|
Total sunshine duration (h) |
648.0 |
421.9 |
|
Wintering to podding |
Average temperature (℃) |
8.1 |
9.0 |
|
Precipitation (mm) |
128.0 |
264.5 |
|
Total sunshine duration (h) |
600.5 |
487.7 |
|
Podding to ripening |
Average temperature (℃) |
19.6 |
19.1 |
|
Precipitation (mm) |
156.5 |
103.5 |
|
Total sunshine duration (h) |
264.8 |
340.2 |
|
The whole growth stage |
Average temperature (℃) |
9.9 |
10.4 |
|
Precipitation (mm) |
382.5 |
516.5 |
|
Total sunshine duration (h) |
1513.3 |
1249.8 |
Table 2Silique shattering resistance index (SSRI) of different accession in 2023-2024.
Table 2
|
Group |
Accession name |
2023 |
2024 |
|
A |
Jungmo7002 |
|
0.42ef |
|
0.27cd |
|
Jungmo7003 |
|
0.48de |
|
0.29cd |
|
Yeongsan |
|
0.4fg |
|
0.12gh |
|
Tammi |
|
0.53cd |
|
0.18ef |
|
Naehan |
|
0.54cd |
|
0.29cd |
|
Mokpo133 |
|
0.14ij |
|
0.05ij |
|
Mokpo134 |
|
0.52cd |
|
0.26cd |
|
IT031247 |
|
0.23hi |
|
0.03j |
|
IT031264 |
|
0.21hi |
|
0.15fg |
|
IT031274 |
|
0.75b |
|
0.38c |
|
IT031276 |
|
0.15ij |
|
0.08hi |
|
IT031294 |
|
0.88a |
|
0.58b |
|
IT031325 |
|
0.84ab |
|
0.23de |
|
IT031353 |
|
0.40fg |
|
0.19ef |
|
IT031359 |
|
0.62c |
|
0.22de |
|
IT031375 |
|
0.88a |
|
0.85a |
|
B |
Jungmo7001 |
|
0.23hi |
|
0.27cd |
|
Yuryeo |
|
0.44de |
|
0.63b |
|
Halla |
|
0.31gh |
|
0.34cd |
|
Tamra |
|
0.3gh |
|
0.05ij |
|
IT031275 |
|
0.22hi |
|
0.32cd |
|
IT031277 |
|
0.05j |
|
0.33cd |
|
IT031283 |
|
0.13ij |
|
0.18ef |
|
IT031292 |
|
0.28h |
|
0.37c |
|
IT031319 |
|
0.08j |
|
0.09hi |
|
IT031377 |
|
0.08j |
|
0.18ef |
|
IT031386 |
|
0.08j |
|
0.11gh |
|
IT031390 |
|
0.05j |
|
0.34cd |
|
Mean |
|
0.37 |
|
0.26 |
|
CV% |
70 |
|
70 |
|
Table 3Agronomic traits of siliques of accessions in 2023-2024.
Table 3
|
Silique trait |
Min |
|
Max |
|
Mean |
|
Median |
|
Variance coefficient (%) |
|
|
|
|
|
|
2023 |
2024 |
2023 |
2024 |
2023 |
2024 |
2023 |
2024 |
2023 |
2024 |
|
Silique length (mm) |
66.20 |
65.99 |
|
114.25 |
106.12 |
|
86.30 |
86.70 |
|
85.96 |
86.59 |
|
10.64 |
8.69 |
|
Pedicel length (mm) |
11.25 |
12.55 |
|
30.80 |
34.20 |
|
19.16 |
19.01 |
|
19.09 |
18.72 |
|
16.06 |
14.98 |
|
Beak length (mm) |
9.39 |
6.72 |
|
21.48 |
18.53 |
|
14.12 |
12.17 |
|
14.02 |
12.05 |
|
16.31 |
16.26 |
|
Silique thickness (mm) |
2.56 |
2.46 |
|
3.86 |
3.62 |
|
3.15 |
3.10 |
|
3.14 |
3.08 |
|
8.77 |
7.70 |
|
20-silique weight (g) |
1.83 |
2.02 |
|
5.23 |
4.88 |
|
3.20 |
3.35 |
|
3.15 |
3.37 |
|
19.59 |
13.86 |
|
20-silique seed weight (g) |
0.69 |
0.15 |
|
2.74 |
2.56 |
|
1.70 |
1.70 |
|
1.68 |
1.71 |
|
21.42 |
20.43 |
|
silique wall thickness (mm) |
1.78 |
1.96 |
|
3.12 |
3.43 |
|
2.41 |
2.72 |
|
2.40 |
2.73 |
|
11.06 |
11.64 |
|
20-silique wall weight (g) |
0.52 |
0.62 |
|
1.63 |
1.65 |
|
0.97 |
1.06 |
|
0.96 |
1.06 |
|
22.08 |
17.06 |
|
Septum thickness (mm) |
0.15 |
0.17 |
|
0.36 |
0.52 |
|
0.24 |
0.24 |
|
0.24 |
0.24 |
|
12.52 |
20.66 |
|
Pedicel-septum juction thickness (mm) |
1.09 |
1.02 |
|
1.99 |
2.59 |
|
1.41 |
1.33 |
|
1.40 |
1.32 |
|
11.06 |
11.39 |
Table 4Comparison of silique shatter resistance index (SSRI) and agronomic traits in different group in 2023-2024.
Table 4
|
Group |
Year |
SSRI |
SL |
PL |
20-SW |
20-SSW |
20-SWW |
PST |
|
A |
2023 |
0.50 |
a |
88.18 |
ns |
19.84 |
ns |
3.51 |
ns |
1.87 |
a |
1.07 |
ns |
1.42 |
ns |
|
2024 |
0.27 |
b |
86.56 |
|
19.41 |
|
3.25 |
|
1.60 |
b |
1.06 |
|
1.34 |
|
|
B |
2023 |
0.19 |
ns |
83.37 |
ns |
19.20 |
ns |
3.14 |
ns |
1.74 |
ns |
0.94 |
b |
1.34 |
ns |
|
2024 |
0.25 |
|
85.94 |
|
19.03 |
|
3.39 |
|
1.56 |
|
1.22 |
a |
1.32 |
|
References
- 1. Braatz J, Harloff HJ, Emrani N, Elisha C, Heepe L, Gorb SN, Jung C. 2018. The effect of INDEHISCENT point mutations on silique shatter resistance in oilseed rape (Brassica napus). Theor Appl Genet 131: 959-971.
- 2. Bruce DM, Farrent JW, Morgan CL, Child RD. 2002. Determining the oilseed rape pod strength needed to reduce seed loss due to pod shatter. Biosyst Eng 81: 179-184.
- 3. Hu Z, Hua W, Huang S, Yang H, Zhan G, Wang X, Liu G, Wang H. 2012. Discovery of pod shatter-resistant associated SNPs by deep sequencing of a representative library followed by bulk segregant analysis in rapeseed. PLOS ONE 7: e34253.
- 4. Hu Z, Yang H, Zhang L, Wang X, Liu G, Wang H, Hua W. 2015. A large replum-valve joint area is associated with increased resistance to pod shattering in rapeseed. J Plant Res 128: 813-819.
- 5. Jeong J, An DH, Cha YL, Choi JB, Kim SY, Kim KS. 2024. 'Yuryeo': A rapeseed with high oleic acid, resistant to Sclerotinia stem rot and lodging. Korean J Breed Sci 56: 53-61.
- 6. Kuai J, Sun Y, Liu T, Zhang P, Zhou M, Wu J, Zhou G. 2016. Physiological mechanisms behind differences in pod shattering resistance in rapeseed (Brassica napus L.) varieties. PLOS ONE 11: e0157341.
- 7. Liu J, Mei DS, Li YC, Cui JC, Wang H, Peng PF, Fu L, Hu Q. 2013. Combining ability and breeding potential of rapeseed elite lines for pod shatter resistance. J Integr Agric 12: 552-555.
- 8. Liu J, Wang J, Wang H, Wang W, Zhou R, Mei D, Cheng H, Yang J, Raman H, Hu Q. 2016. Multigenic control of pod shattering resistance in Chinese rapeseed germplasm revealed by genome-wide association and linkage analyses. Front Plant Sci 7: 1058
- 9. Meakin PJ, Roberts JA. 1990a. Dehiscence of fruit in oilseed rape (Brassica napus L.): I. Anatomy of pod dehiscence. J Exp Bot 41: 995-1002.
- 10. Meakin PJ, Roberts JA. 1990b. Dehiscence of fruit in oilseed rape (Brassica napus L.): II. The role of cell wall degrading enzymes and ethylene. J Exp Bot 41: 1003-1011.
- 11. Morgan CL, Bruce DM, Child R, Ladbrooke ZL, Arthur AE. 1998. Genetic variation for pod shatter resistance among lines of oilseed rape developed from synthetic B. napus. Field Crops Res 58: 153-165.
- 12. Nath UK, Kim H, Khatun K, Park J, Kang K, Nou I. 2016. Modification of fatty acid profiles of rapeseed (Brassica napus L.) oil for using as food, industrial feed-stock and biodiesel. Plant Breed Biotech 4: 123-134.
- 13. Piao X, Choi SY, Kim YH, Lee YH, Kim KS, Jang YS, So YS, Kim HS. 2013. Variety×Location interaction on oil, fatty acids, tocopherols and phytosterols in Korean rapeseed (Brassica napus L.). Plant Breed Biotech 1: 91-101.
- 14. Price JS, Hobson RN, Neale MA, Bruce DM. 1996. Seed losses in commercial harvesting of oilseed rape. J Agric Eng Res 65: 183-191.
- 15. Qing Y, Li Y, Xu L, Ma Z, Tan X, Wang Z. 2021. Oilseed rape (Brassica napus L.) pod shatter resistance and its relationship with whole plant and pod characteristics. Ind Crops Prod 166: 113459.
- 16. RDA, Rural Development Administration.2012. Criteria for agricultural research and investigation.
- 17. Shen J, Liu Y, Wang X, Bai J, Lin L, Luo F, Zhong H. 2023. A comprehensive review of health-benefiting components in rapeseed oil. Nutrients 15: 999
- 18. Wang R, Ripley VL, Rakow G. 2007. Pod shatter resistance evaluation in cultivars and breeding lines of Brassica napus, B. juncea and Sinapis alba. Plant Breed 126: 588-595.
- 19. Weymann W, Bottcher U, Sieling K, Kage H. 2015. Effects of weather conditions during different growth phases on yield formation of winter oilseed rape. Field Crops Res 173: 41-48.