적요
Starch is the most important carbohydrate in the human and animal diets and it also has numerous industrial applications. Starch is widely used as a key material for processed foods such as bread, pancakes, cereals, noodles, pasta, porridge and tortilla. Increasing the starch content in food crops is one of the important targets for crop breeding. We used a starch-iodine test to identify varieties with high starch content in 2354 soybean germplasm collection and found a total of 126 germplasm that showed a strongly positive reaction. We quantified starch content from highly stained 7 soybean accessions by standard method (glucose-oxidase). As results, starch contents of 7 soybean germplasm were 2.81~4.55%, whereas the weakly stained controls showed low starch content with less than 1%. We also measured the contents of protein, fatty acid, water potential and free sugar in 7 soybean germplasm with high starch content. High starch germplasm showed lower protein content, while low starch germplasm showed higher protein content. Our results suggest that the increasing of starch content could affect protein level in soybean seed.
서 언
콩(
Glycine max (L.) Merr.)은 단백질(43~48%), 지질(18 ~21%) 함량이 높고 필수 아미노산과 필수 지방산이 풍부하 다. 또한 탄수화물인 sucrose (4.9~6.8%), raffinose (0.8~ 1.2%)와 stachyose (3.5~4.3%)을 함유하고 있다(
Hildebrand et al. 1981,
Wilcox & Shibles 2001). 이러한 영양성분 이외 에도 isoflavones, oligosaccharides, saponins, phytic acids, trypsin inhibitors 등 생리활성물질을 다량 보유하고 있다. 이 에 우리나라를 비롯한 동양에서는 중요한 단백질과 지질의 공급원으로 이용되고 있다. 콩은 우수한 단백질 공급원으로서 국내에서는 된장, 간장, 두부, 콩나물, 밥밑콩 등 다양한 전통 식품의 가공, 조리에 이용되는 영양식품의 원료로 이용되고 있다. 특히 콩 단백질은 양질의 식물성 단백질로서 가공기술 의 발전에 따라 다른 식품에 첨가되는 단백질 보강 재료로도 이용되고 있다. 최근 콩 단백질의 식품적 우수성이 인정되면 서 세계적으로 수요가 증가하고 있는 실정이다.
전분은 동물에게 중요한 탄수화물 에너지원으로 사용되며, 다양한 산업적 가공성을 가지고 있다. 콩은 높은 단백질과 지 방함량을 가지고 있지만, 상대적으로 낮은 전분함량을 나타낸 다. 따라서, 콩 종자에서 낮은 전분함량은 콩의 산업적 이용성 을 크게 저해하고 있다. 녹두, 팥, 완두와 같은 두과 작물은 약 34~55%의 높은 전분 함량을 나타내지만, 이에 비해 콩은 1% 미만의 전분을 함유하고 있는 것으로 알려져 있다 (
Wilson et al. 1978,
Jeong et al. 2010). 수확하기 20일 전까지의 등 숙기간 동안 콩의 전분함량은 10~20%로 높게 축적이 된다 (
Yazdi-Samadi et al. 1977). 그러나, 성숙된 콩 종자의 전분 함량은 0.19~0.91%로 상당히 낮게 나타난다(
Wilson et al. 1978). 이러한 성숙시기에 따른 전분함량 감소는 가인산분해 (phosphorolysis)의 증가에 따른 전분분해 증가에 기인하는 것으로 유추되어져 있다(
Monma et al. 1991). 그러나, 전분 의 분해에 의해 생성된 탄소원이 단백질과 지방의 합성에 어 떻게 이용되는지에 대해서는 거의 연구가 되어있지 않다. 최 근의 연구 결과들은 전분의 특성이 콩의 조성 성분에 영향을 줄 수 있음을 시사해주고 있다. Stevenson et al. (
2006)은 다 른 지방 조성을 보이는 콩 픔종은 다른 전분 구조를 보인다고 보고하였다. 저 리놀레닉산(low-linolenic acid) 콩 품종의 전 분은 고단백 콩 품종 이나 lypoxygrease-free 콩 품종의 전분 보다 상대적으로 높은 아밀로즈(amylase) 함량을 포함하고 있었다. 이 연구는 성숙된 콩 종자가 아니라, 등숙기간 중의 미성숙된 종자를 이용하였다. 따라서, 미성숙된 콩 종자의 아 밀로즈 함량이 성숙된 종자의 지방조성에 영향을 주는지에 대해서는 더 깊은 연구가 필요하다.
식품에 전분을 사용하는 이유는 영양적 목적보다는 기능성 을 개선하기 위함인데 색깔, 조직, 씹음촉각, 식품성분의 유리 방지, 풍미 등의 개선에 주로 사용하고 있다. Urszula et al. (
2010)의 연구에 따르면, 감자 전분, 옥수수 전분, 콩 전분으 로 빵을 제조했을 때 콩 전분 빵은 향상된 화학적 조성과 품 질을 지니고 있어 빵의 크럼 굳기를 감소시키고 탄력성을 높 여주어 완성된 빵에 더 좋은 질감을 부여했다고 한다. 따라서, 고 전분함량 콩 종자의 사용은 콩 식품의 가공성에 많은 영향 을 줄 수 있을 것이라 여겨진다. 그러나, 현재까지 콩 등숙기 동안 전분의 합성과 분해에 대한 명확한 분자적 기작은 밝혀 져 있지 않다. 또한, 전분함량에 영향을 미치는 유전적 요인이 나 유전 양상에 대한 연구는 거의 되어있지 않은 실정이다.
국내 육성 콩 주요품종 및 보존 유전자원을 대상으로 전분 함량을 포괄적으로 검토한 사례가 전무한 실정이다. 따라서, 본 연구는 국내 콩 주요 품종 및 보존 유전자원의 전분함량과 변이 양상을 분석하여 고 전분 함유 유용 유전자원 선발 및 콩 우량품종 육성의 기초자료로 활용하는데 있다. 더 나아가 콩의 산업적 활용도 증진을 위한 국내 자원의 이용성 증진 및 콩 유전자원 연구에 기초를 제공하고자 하였다.
재료 및 방법
실험재료
본 실험에 사용된 콩 주요품종 및 보존 유전자원 등 2354 점은 농촌진흥청 국립식량과학원 기능성작물부 대구시험지에 서 2012년 6중순에 파종하여 재배한 후 수확하였다.
고 전분함량 콩 유전자원의 선발
2354점의 유전자원 중 전분함량이 높은 유전자원을 1차로 선발하기 위해 요오드-전분 반응(iodine-starch reaction)으로 분석하였다. 콩 종실을 단면으로 절단한 후 요오드 용액(2% I2와 0.5% KI) 약 20 μl를 첨가하였다. 약 10분 동안 실온에 서 반응한 후 염색되는 정도에 따라 등급을 ‘1’에서 ‘4’로 분 류를 하였다. ‘1’은 무반응, ‘2’는 연한 청남색, ‘3’은 청남색, ‘4’는 진한 청남색을 나타낸다. Vibrating sample mill TI-100 (CMT, Japan)를 이용하여 분쇄하여 전분, 조단백질, 조지방, 수분, 유리당함량 분석에 사용하였다.
전분함량 분석
총 전분함양은 total starch kit (Megazyme International Ireland Ltd., Wicklow, Ireland)를 이용하여 AOAC (
McCleary et al. 1997,
Lee et al. 2011) 방법에 의하여 측정하였다. 시 료 약 0.1 g에 80% 에탄올 0.2 mL을 가하고, 열에 안정한 α-amylase 3.0 mL을 가하여 교반 후 끓는 물에서 6분간 반응시 켰다. 실온에서 약 50°C로 냉각 시킨 후 amyloglucosidase 0.1 mL을 가하여 교반 후 50°C에서 30분간 반응시켰다. 멸균된 물로 시료 용액을 10 mL로 맞춘 후 3,000 rpm에서 10분간 원 심 분리하였다. 상등액 0.1 mL을 취하여 GOPOD reagent 3.0 mL을 가하고 50°C에서 20분간 발색시킨 후 UV/VIS Spectrophotometer (Libra S35, Biochrom Ltd., Cambridge, UK)를 이용하여 흡광도(510 nm)를 측정하여 아래의 식 (
1) 으로 총 전분함량을 측정하였다.
ΔA : Absorbance
F : 100 μg of D-glucose / absorbance for 100 μg of D-glucose
FV : Final Voume
W : Weight of sample (mg)
조단백질 함량 분석
단백질의 함량 분석시 0.2 g의 분쇄시료를 사용하였으며 Rapid N cube (Elementar Analysensysteme GmbH, Germany) 로 분석하였다. 콩의 질소 환원계수 6.25를 곱하여 조단백질 의 함량을 계산하였다.
조지방 함량 분석
조지방은 자동 속시렛 추출장치(Extraction unit E-816, BUCHI)를 이용하여 추출하였다. 콩 분쇄시료 2.0 g에
n-hexane 200 mL을 추출용매로 첨가하여 2시간 동안 추출하였고, 10 5°C에서 1시간 동안 건조한 뒤 방냉하고, 추출된 조지방의 무 게를 측정하여 조지방 함량으로 계산하였다(
Choung 2006).
수분함량은 분쇄시료 약 1.0 g을 자동 수분 측정기 (MX50, AND)를 이용하여 측정하였다.
유리당 분석
유리당 분석은 분쇄시료 1.0 g에 10 mL 80% 에탄올 수 용액으로 상온에서 24시간 진탕 추출하였다. 추출된 시료는 3,000 rpm으로 원심분리 후 상등액을 취하여 0.45 μm 필터 로 여과하여 2배 희석한 시료를 HPLC로 분석하였다. 분석 에 사용된 HPLC는 Agillent 1100 series (Wilmington, DE) 를 이용하였고, 분석용 칼럼은 Sugar-Pak Ⅰ(Waters Co. Milford, MA)이며, 용매는 초순수 증류수를 이용하여 분당 유속을 0.5 mL/min하고, 칼럼온도를 80°C로 유지하며, 시료 주입량을 20 μL로 조절하여 분석하였다. 분석에 사용된 굴절 율 검출기는 Shodex RI-101 (Japan) 검출기를 사용하였고, 분석에 사용된 당 표준품은 모두 Sigma-Aldrich (St. Louis, Mo, USA)에서 구입하여 이용하였다.
결과 및 고찰
고 전분 함량을 나타내는 콩 품종을 선발하기 위해서, 콩 유전자원 2354점에 대해서 요오드-전분 반응을 실시하여 착 색 정도를 관찰하였다. 착색 정도에 따라 1에서 4등급으로 분 류를 하였으며(Fig.
1A), 대부분의 콩 유전자원이 2~3등급 으로 분류되었다. 가장 강한 착색 반응을 보인 4등급에 속하 는 유전자원은 126점 이었다(Fig.
1B). 전분함량의 정량적 분 석을 위해 4등급으로 분류된 콩 유전자원들 중 가장 착색 반 응이 높은 7점, 1~2등급으로 분류된 콩 유전자원 3점에 대해 total starch kit (Megazyme International Ireland Ltd., Wicklow, Ireland)을 이용하여 전분함량을 측정하였다. 그 결과, 1, 2등 급으로 분류된 유전자원 Daewang, Soheonje, Jyuiku #109 는 각각 0.57, 0.45, 0.54%의 낮은 전분함량을 나타내었다 (Table
1). 반면, 4등급으로 분류된 유전자원은 2.81~4.55% 의 상대적으로 높은 전분함량을 나타내었다. 특히 Mansu, Gwangjusetae, Daeha의 전분함량은 4%이상으로 나타나 유 전자원 대량검정에 요오드-전분 반응법을 이용하는 것이 가 능할 것으로 판단되었다.
Fig. 1.Screening of soybean germplasm with high starch contents by iodine-starch reaction. A, Histochemical analysis in matured seeds of soybean germplasm. B, Histograms of reaction levels in 2354 soybean germplasm.

Table 1Quantification of starch content of soybean germplasm
Table 1
|
Germplas |
grade |
Starch (%) |
|
|
Taekwang |
4 |
3.48 ± 0.46 |
|
Mansu |
4 |
4.03 ± 0.23 |
|
Sobaegnamul |
4 |
3.20 ± 0.11 |
|
Hoban |
4 |
3.60 ± 0.04 |
|
Gwangjusetae |
4 |
4.26 ± 0.14 |
|
Daeha |
4 |
4.55 ± 0.18 |
|
1340 |
4 |
2.81 ± 0.11 |
|
Daewang |
1 |
0.57 ± 0.02 |
|
Soheonje |
1 |
0.45 ± 0.00 |
|
Jyuiku #109 |
1 |
0.54 ± 0.01 |
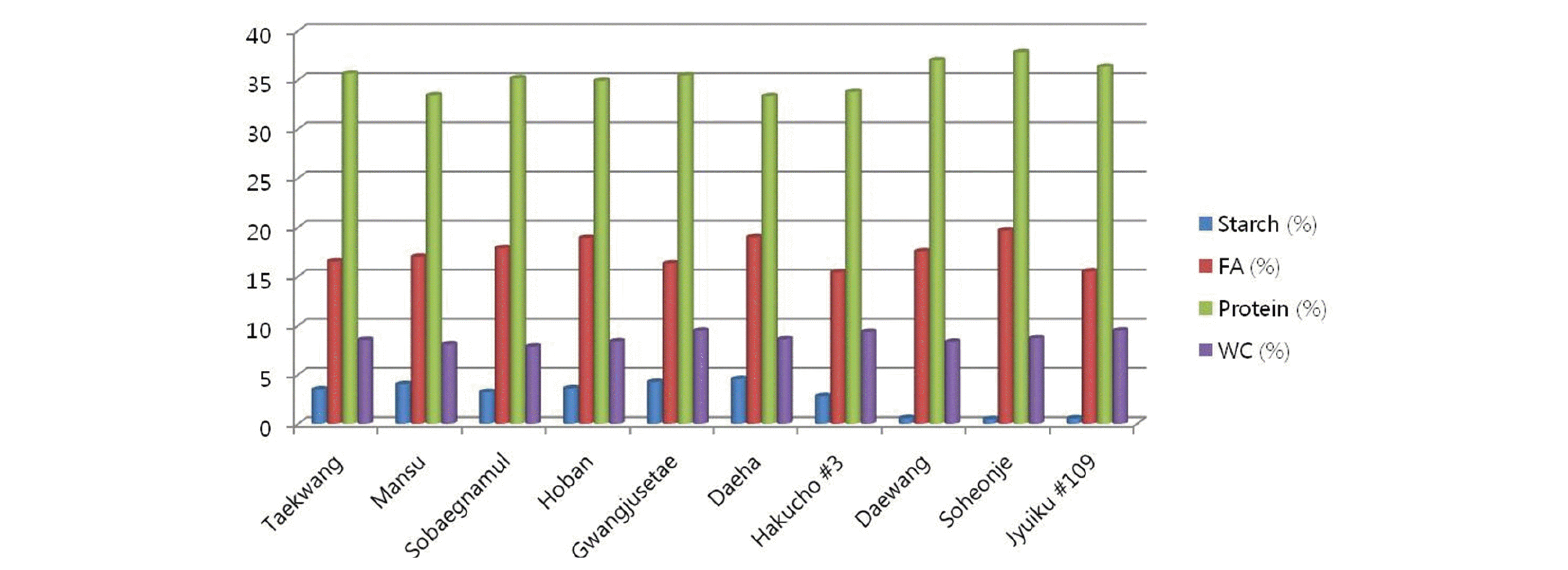
각 유전자원에서 전분함량의 차이가 다른 구성 성분의 함 량에 영향을 미치는지를 알아보기 위해, 앞의 유전자원 10점 에 대해서 조단백질함량, 조지방함량, 수분함량을 각각 측정 하였다. 조사된 유전자원의 조단백질 함량은 33.33~37.79% 를 나타내었다(Fig.
2). 전분함량이 높은 4등급 유전자원의 조단백질함량은 33.33~35.63%로 전체적으로 낮은 경향성을 나타내었다. 반면, 전분함량이 낮은 1,2등급 유전자원의 조단 백질 함량은 36.32~36.98%로 상대적으로 높게 나타났다. 따라서, 이 결과는 전분함량의 변화는 조단백질 함량 변화에 영향을 미칠 수 있음을 제시해주고 있다. 즉, 전분 분해 후 그 산물들은 단백질 합성대사에 사용되며, 콩 종자의 단백질 함 량을 증가시키는 것으로 추측된다. 전분의 분해에 의해 세포 내 탄소의 비율이 증가 되며, 이는 단백질 합성 대사시 탄소 원으로 사용 되어지는 것으로 알려져 있다. 그리고, 전분 분해 에 의해 생성되는 포도당은 단백질 합성을 포함한 세포내 대 사 작용에 필요한 에너지원으로 사용된다(
Smith et al. 2005). 최근의 보고에 의하면, 등숙기의 콩은 약 10~20%의 전분을 포함하지만, 완전 등숙된 콩 종자는 약 1% 미만의 전분을 포 함하는 것으로 알려져 있다. 따라서, 전분의 분해는 단백질 합 성 대사에 중요한 역할을 함을 알 수 있다. 조지방 함량의 범위 는 15.40~19.66% 였으며, 수분함량의 범위는 7.85~9.48% 으로 나타났다(Fig.
2). 전분함량과 비교 분석했을 때, 전분함 량의 차이가 조지방과 수분함량의 변화에 영향을 미치지 않 는 것으로 유추된다. 각 유전자원에서 조지방과 수분함량의 차이는 유전자원 개체별 차이에 기인하는 것으로 사료된다. 전분의 분해에 의해서 당의 생성이 증가되기 때문에, 유전자 원에서 유리당의 변화가 있는지를 분석하였다. 표2에서 보는 바와 같이 유리당은 각 유전자원 개체에서 큰 변이차를 보였 다. 이는 전분변화의 차이에 의한 영향 보다는 개체간의 유전 적 요인에 의한 것이라 유추된다. (Table
2).
Fig. 2.Contents of starch, protein, fatty acid (FA) and water content (WC) of soybean.germplasm.
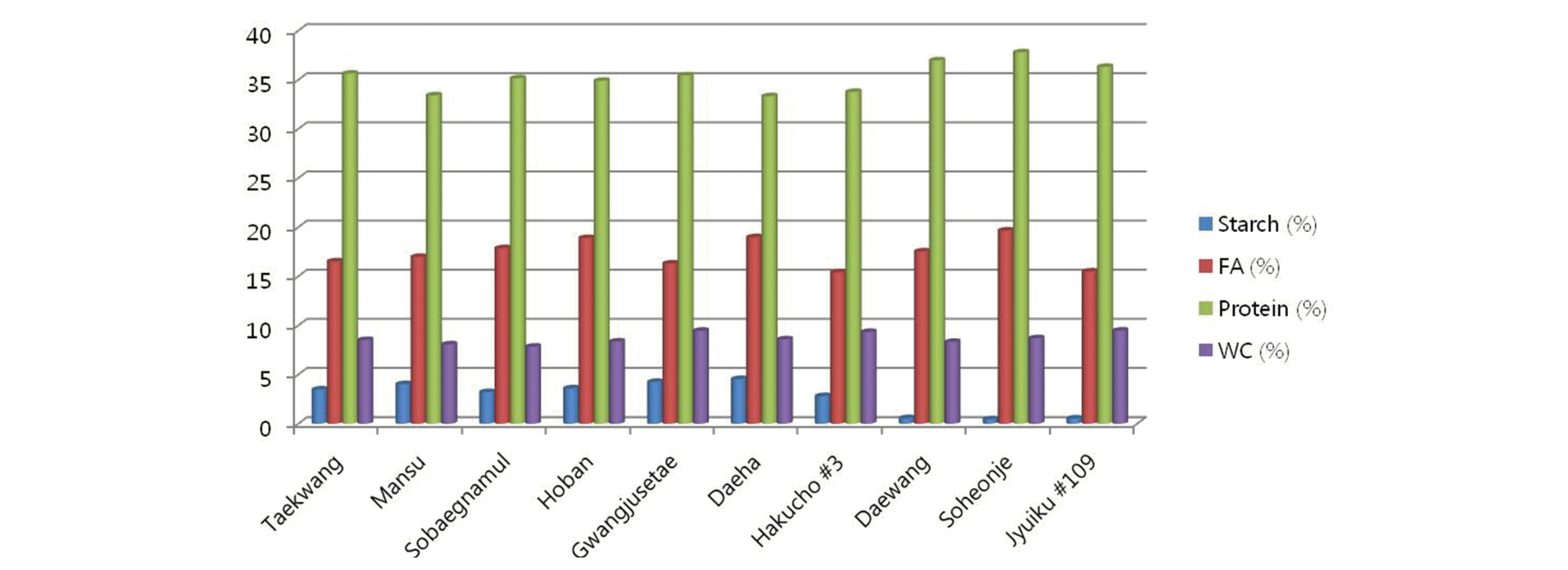
Table 2Free sugar contents of high starch germplasm
Table 2
|
Sample |
Stachyose |
Raffinose |
Sucrose |
Glucose |
Total |
|
|
Taekwang |
24.1 |
0.2 |
82.0 |
2.1 |
108.4 |
|
Mansu |
4.8 |
4.0 |
84.1 |
1.5 |
94.3 |
|
Sobaegnamul |
22.9 |
0.2 |
71.3 |
- |
94.3 |
|
Hoban |
19.9 |
0.2 |
67.0 |
1.0 |
88.2 |
|
Gwangjusetae |
10.9 |
2.0 |
71.1 |
2.2 |
86.2 |
|
Daeha |
9.7 |
2.5 |
80.2 |
- |
92.4 |
|
1340 |
13.5 |
3.5 |
40.3 |
0.3 |
57.6 |
|
Daewang |
21.8 |
0.2 |
62.6 |
- |
84.6 |
|
Soheonje |
5.3 |
1.7 |
12.9 |
19.5 |
39.4 |
|
Jyuiku #109 |
34.9 |
0.2 |
63.8 |
2.7 |
101.6 |
이상의 결과에서 보는 바와 같이, 본 연구에서 전분함량이 높은 콩 유전자원 선발 실험을 통해 126점의 품종을 선발하 였다. 그 중 7점에 대해 정량적 분석을 실시한 결과 3점 (Mansu, Gwangjusetae, Daeha)이 4% 이상의 높은 함량을 나타내는 품종으로 선발 되었다. 그리고, 콩 구성 성분 분석 실험 결과는 전분 함량의 차이는 단백질 함량의 차이에 영향 을 줄 수 있음을 보여주고 있다. 현재까지, 콩에서 전분의 함 량을 결정하는 유전적 요인에 대한 연구가 미흡한 상황이다. 따라서, 선발된 전분함량이 높은 품종들은 콩에서 전분 함량 조절 기작을 구명하는데 중요한 재료로 사용될 것이라 기대 한다. 최근 보고에 의하면, 온도의 증가는 땅콩에서 전분 분해 를 저해한다고 알려져 있다(
Golombek et al. 1995) 그리고, 콩의 전분함량은 등숙기별로 큰 변이를 보인다. 따라서, 전분 함량이 등숙기 동안 온도, 일조량 등과 같은 환경적 요인에 의해 영향을 받는지에 대한 연구도 수행될 필요가 있다. Monma et al. (
1991)은 등숙기 동안 증가된 phosphorylase 와 phytase의 활성이 전분분해에 관여한다고 보고 하였다. 이 연구 결과와 같이 전분 분해 효소의 활성 조절 기작에 대한 연구는 콩에서 전분 함량을 조절하는 유전적 인자를 구명하 는데 중요한 역할을 할 것이라 생각 된다. 따라서, 이러한 전 분 함량이 높은 품종을 이용한 전분함량 조절 기작의 구명은 전분 함량이 높은 새로운 기능성 콩 품종 개발에 기여할 수 있을 것이라 기대된다.
적 요
본 연구는 고 전분 함유 유용 유전자원 선발 및 콩 우량품 종 육성의 기초자료로 활용하고자 국내 콩 주요 품종 및 보존 유전자원의 전분함량과 변이 양상을 분석하였으며 그 결과는 다음과 같다.
총 2354점 국내 콩 주요 품종 및 보존 유전자원에 대한 요 오드-전분 반응에 의한 착색 정도에 따라 1에서 4등급으로 분류를 하였으며, 대부분의 콩 유전자원이 2~3등급으로 분류되었고, 가장 강한 착색 반응을 보인 4등급으로 126점 의 유전자원이 선발되었다
전분함량의 정량적 분석을 수행한 결과, 1, 2등급으로 분 류된 유전자원 Daewang, Soheonje, Jyuiku #109는 각각 0.57, 0.45, 0.54%의 낮은 전분함량을 나타내었다. 반면, 4등급으로 분류된 유전자원은 2.81~4.55%의 상대적으로 높은 전분함량을 보여 요오드-전분 반응법은 유전자원의 대량검정에 유용하게 이용할 수 있을 것으로 판단되었다.
각 유전자원에서 전분함량의 차이가 다른 구성 성분의 함 량에 영향을 미치는지를 알아보기 위해, 조단백질함량, 조 지방함량, 수분함량을 각각 측정하였다. 전분함량의 변화 는 조단백질 함량 변화에 영향을 미치며, 조지방과 수분함 량의 변화에 영향이 없음을 확인하였다.
사 사
본 연구는 농촌진흥청 연구사업(ATIS 주관과제번호: PJ009291) 의 지원에 의해 이루어진 것임.
References
- 1. Choung MG. Variation of oil contents andfatty acid compositions in Korean Soybean germplasms. Kor. J. Crop Sci 2006. 51: S. 139-149.
- 2. Golombek SD, Sridhar R, Singh U. Effect of soil temperature on the seed composition of three Spanish cultivars of groundnut (Arachis hypogaea L.). J. Agric. Food Chem 1995. 43: 2067-2070.
- 3. Hildebrand DF, Hymowitz T. Role of b-amylase in starch metabolism during soybean seed development and germination. Physilogia Plantarum 1981. 53: 429-434.
- 4. Jeong W, Harada K, Yamada T, Abe J, Kitamura K. Establishment of new method for analysis of starch contents and varietal differences in soybean seeds. Korean J. Breed. Sci 2010. 60: 160-163.
- 5. Lee SB, Lee JH, Shin YS, Lee KS, Hwang HG, Jeong OY, Yang CI, Choi YH, Yang SJ, Jeon YH, Hong HC, Kim HY, Cho YC, Lee JH, Yea JD, Oh MK, Kim MK, Kim YG, Jeong KH, Lee YT. A medium-maturing, high non-dietary starch, specialty rice cultivar ‘Goami 3’. Korean J. Breed. Sci 2011. 43: 595-599.
- 6. McCleary BV, Gibson TS, Mugford DC. Measurement of total starch in cereal products by amyloglucosidase - α-amylase method: Collaborative study. J. AOAC Int 1997. 80: 571-579.
- 7. Monma M, Sugimoto T, Kawamura Y, Saio K. Starch breakdown in developing soybean seeds. Agric. Biol. Chem 1991. 55: 67-71.
- 8. Urszula K, Cristina MR, Jadwiga S, Maria SŚ. Bean starch as ingredient for gluten free bread. J. Food Process. Preserv 2010. 34: 501-518.
- 9. Smith AM, Zeeman SC, Smith SM. Starch degradation. Annu. Rev. Plant Biol 2005. 56: 73-98.
- 10. Stevenson DG, Doorenbos RK, Jane J, Inglett GE. Structures and functional properties of starch from seeds of three soybean (Glycine max (L.) Merr.) varieties. Starch/Starke 2006. 58: 509-519.
- 11. Yazdi-Samadi B, Rinnie RW, Sief RD. Components of developing soybean seeds: oil protein, sugars, starch, organic acids, and amino acids. Agronomy J 1977. 69: 481-486.
- 12. Wilcox JR, Shibles RM. Interrelationships among seed quality attributes in soybean. Crop Science 2001. 41: 11-14.
- 13. Wilson LA, Birmingham VA, Moon DP, Snyder HE. Isolation and characterization of starch from mature oybeans. Cereal Chemistry 1978. 55: 661-670.