Abstract
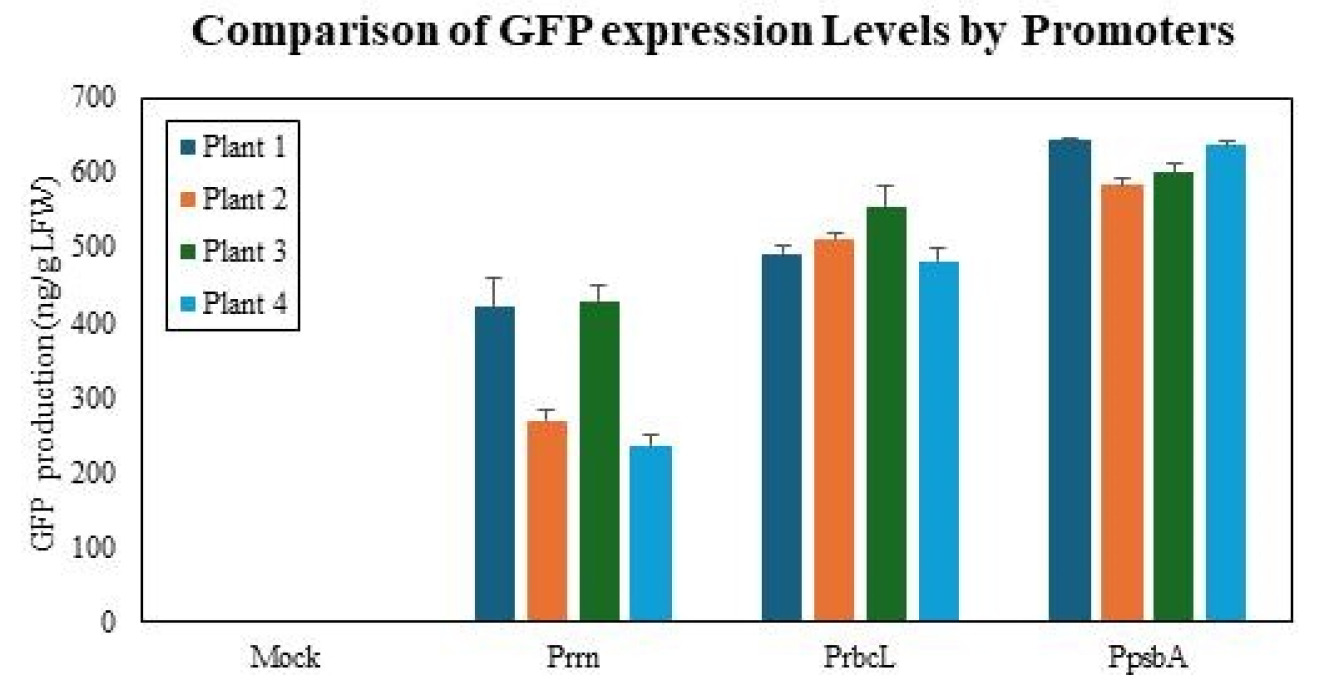
Plant-based production of recombinant proteins has emerged as an efficient and cost-effective alternative to microbial fermentation and mammalian cell culture systems. Chloroplasts harbor high plasmid copy numbers and can be stably transformed, making them efficient platforms for protein production. In the present study, we used green fluorescent protein (GFP) as a reporter to compare the three major chloroplast promoters (rrn, psbA, and rbcL) involved in protein production in Nicotiana tabacum cv. “Petit Havana.” Three chloroplast transformation vectors were constructed, each regulated by a different promoter, and the transformation was performed via biolistic particle bombardment. Transformants were selected based on spectinomycin resistance and were confirmed by PCR. Among the three promoters, psbA showed the highest transformation efficiency and protein expression levels. Reverse transcription quantitative PCR showed that the mRNA levels (relative to Actin) for psbA (218.21±19.64) were nearly twice that of rbcL (126.60±8.78), and five times that of rrn (43.27±1.57). This transcriptional hierarchy was also observed at protein level. Immunoblotting showed the GFP levels (relative to psbA) were: psbA (1.00), rbcL (0.87), and rrn (0.77), whereas quantification through ELISA revealed relative GFP concentrations of: 616.2±28.7 ng/g LFW for psbA, 510.3±32.4 ng/g LFW for rbcL, and 338.9±100.2 ng/g ng/g LFW for rrn. These quantitative results demonstrate the importance of promoter selection for efficient expression of recombinant proteins in chloroplasts and show that the psbA promoter is suitable for high-efficiency chloroplast expression systems, providing a foundation for advancing plant-based molecular farming.
-
Keywords: chloroplast transformation; recombinant protein; promoter; Nicotiana tabacum
Introduction
The production of recombinant proteins in plants has become a compelling and cost-effective alternative to traditional microbial fermentation and mammalian cell culture systems. Plants can be used to generate many valuable recombinant proteins, including pharmaceuticals, industrial enzymes, and vaccine antigens. They also offer distinct advantages as platforms for recombinant protein production, particularly in terms of cost-effectiveness, scalability, and rapid production (
Feng et al. 2022,
Lee et al. 2023). Plants can be cultivated on an agricultural scale for mass production, reducing production costs to only 2-10% of the costs for traditional microbial culture and 0.1% of costs for mammalian cell culture systems (
Giddings 2001). Because of their fast production rates and high biomass yields, plants are well suited for large-scale production of proteins (
Schillberg et al. 2019). Notably, transient expression systems in plants allow for the rapid generation of target proteins within just a few weeks, a feature that is especially advantageous for urgent applications such as vaccine development (
Venkataraman et al. 2023). These features make plant-based systems a promising and competitive alternative for the current and future production of pharmaceutical and industrial recombinant proteins (
Lee et al. 2023).
For protein production
in planta, the chloroplast has garnered significant attention as an optimal compartment for transgene expression. This is primarily due to its unique genetic architecture and biosynthetic capacity, offering the potential for accumulation of exceptionally high levels of recombinant protein (
Cheon et al. 2018,
Daniell et al. 2016,
Maliga 2004). Chloroplast transformation via the targeted integration of foreign DNA into the chloroplast genome presents several key benefits. First, the high copy number of the chloroplast genome (often thousands of copies per cell) leads to significantly higher protein yields compared to nuclear transformation, with recombinant proteins constituting up to 75% of total soluble protein (TSP) in some cases (
Castiglia et al. 2016,
Oey et al. 2009). Second, foreign genes can be precisely integrated into the chloroplast genome via homologous recombination, allowing for targeted insertion at specific loci (
Bock 2015,
Svab & Maliga 1993). Targeted integration greatly reduces the risk of the position effects and transgene silencing that are frequently encountered in nuclear transformation, thereby ensuring stable and predictable expression of the introduced genes (
Daniell et al. 2016,
Maliga 2004). Third, the prokaryote-like gene expression machinery within chloroplasts, including the presence of operon-like structures, allows for the stable and coordinated expression of multiple genes from a single polycistronic transcript, as with bacterial operons (
Bock 2015). This is particularly advantageous for engineering complex metabolic pathways or for producing multisubunit proteins.
Many studies have demonstrated the successful application of chloroplast transformation over the past decade, including the production of pharmaceutical recombinant proteins. For instance,
Bock et al. (2018) and Su et al. (2023) reviewed progress in engineering chloroplasts for production of vaccines, antibodies, and other therapeutic proteins. A particularly noteworthy application in this area involves the oral delivery of therapeutic proteins. In a groundbreaking study,
Daniell et al. (2020) showed that bioencapsulated forms of angiotensin-converting enzyme 2 and angiotensin-(1-7), produced in lettuce chloroplasts significantly reduced the symptoms of pulmonary hypertension when orally administered to rats.
Wang et al. (2023) achieved high levels of human epidermal growth factor (EGF) by creating a fusion construct with green fluorescent protein (GFP) in tobacco (
Nicotiana tabacum) chloroplasts. The resulting GFP-EGF fusion protein accumulated to 10.21% of TSP, the equivalent to 1.57±0.05 ug/g leaf fresh weight (LFW), representing one of the highest reported accumulation levels for EGF in plant-based systems.
Building upon these therapeutic applications,
Legen et al. (2023) used CHLORORESPIRATORY REDUCTION2-dependent sRNA sequences to drive expression of human papillomavirus (HPV) in tobacco chloroplasts, providing an innovative strategy to produce antigens for vaccine development. They established homoplasmic lines capable of expressing both HPV18 L1 protein and an HPV16B enterotoxin-L1 fusion protein, employing pentatricopeptide repeat protein binding sites as novel expression elements. Demonstrating the industrial potential of this technology for biofuel applications,
Castiglia et al. (2016) successfully used tobacco chloroplasts to produce thermostable cellulolytic enzymes from hyperthermophilic microorganisms. In this study, three thermophilic enzymes were expressed in the chloroplasts of tobacco plants, each showing distinct accumulation levels. Endoglucanase from
Sulfolobus solfataricus accumulated up to 2% of TSP, endo-β-1,4-xylanase from
Alicyclobacillus acidocaldarius reached 35.7% of TSP, and β-glucosidase from
Pyrococcus furiosus exceeded 75% of TSP. These diverse examples underscore the versatility and utility of chloroplast transformation.
A critical determinant for achieving high and predictable transgene expression in chloroplasts is the selection of an appropriate promoter. Several strong plastid promoters have been characterized and widely utilized for
in planta protein production. The
rrn promoter, derived from the ribosomal RNA operon in the tobacco chloroplast genome, is known for its exceptionally strong and largely constitutive activity, enabling robust expression throughout various tissues and developmental stages (
Allison et al. 1993,
Quesada-Vargas et al. 2005). The promoter of
psbA, which encodes the D1 protein of photosystem II, exhibits very high light-inducible activity and has been reported to drive transgene accumulation up to 5-10% of total soluble protein (TSP) in photosynthetically active tissues (
Staub & Maliga 1993,
Staub & Maliga 1994). Similarly, the
rbcL promoter regulates expression of the large subunit of RuBisCO, which can constitute up to 50% of leaf protein, and is another powerful light-regulated promoter frequently employed in chloroplast engineering (
Allison et al. 1993,
Suzuki et al. 2003). Although these promoters have individually demonstrated substantial protein production capacity, their relative efficiencies and regulatory properties differ—for example, the light responsiveness of
psbA, the constitutive activity of
rrn, and the developmental regulation of
rbcL. These distinct features, together with their broad use in plastid transformation, make them particularly appropriate candidates for direct comparative analysis.
In this study, we compared the rrn, psbA, and rbcL promoters to evaluate their efficacy in driving GFP expression within the chloroplasts of tobacco, a model plant widely used for chloroplast transformation. By employing a standardized reporter gene system and consistent transformation and analytical methods, we have provided a direct comparison of the transcriptional strengths and resultant protein accumulation levels driven by these promoters. The insights gained from this research will inform the rational design of chloroplast expression vectors, ultimately contributing to the optimization of recombinant protein production in plants for various biotechnological applications.
Materials and Methods
Plant Materials and Growth Conditions
Tobacco (Nicotiana tabacum cv. Petit Havana) plants were used in this study. Mature seeds were placed in a 1.5-mL microcentrifuge tube and surface sterilized by immersion in 1 mL of 70% ethanol for 1 min, followed by three rinses with sterile distilled water. Subsequently, the seeds were treated with 1 mL of 20% sodium hypochlorite solution for 30 min and rinsed an additional four times with sterile distilled water. The disinfected seeds were then transferred onto Petri dishes lined with three to four layers of sterile filter paper. Air-dried seeds were grown on 0.5×agar-solidified Murashige and Skoog (MS) medium (Duchefa Biochemie, Haarlem, Netherlands, M0222) supplemented with 10 g/L sucrose. Wild-type tobacco plants were cultivated in a growth chamber (Koencon, Korea) at 25℃ with 60% relative humidity, a 16-h photoperiod (110 μmol m-2 s-1), and 400-420 ppm CO2.
Vector Construction and Cloning
Three promoter sequences,
rrn (NCBI accession no. JX235341, nt 5480-5614),
rbcL (NCBI accession no. MT596798, nt 815-927), and
psbA (NCBI accession no. KY426959, nt 6176-6278), were obtained from the NCBI database and synthesized. The synthesized promoters were then inserted into the pBL vector using the restriction enzymes KpnI and XhoI to complete the construction of the recombinant plasmids. All cloning steps were performed using standard molecular biology techniques, and the final construct was confirmed by restriction digestion and DNA sequencing (
Sambrook & Russell 2001).
The pBL Prrn/PrbcL/PpsbA-GFP-TpsbA vectors were transformed into 4-week-old tobacco leaves by particle bombardment. For each bombardment, six or seven fully expanded leaves (approximately 4 cm
2) were placed adaxial side up on antibiotic-free MS medium containing 0.1 μg/mL NAA, 1.0 μg/mL BAP, 1.0 μg/mL thiamine-HCl, and 100 μg/mL inositol. Leaves were bombarded with 0.6-μm gold particles (Bio-Rad, Hercules, CA, USA), which were coated with plastid transformation vectors using a DNA precipitation method with 2.5 M CaCl
2 and 0.1 M spermidine. Bombardments were performed with the PDS-1000/He Biolistic device (Bio-Rad), using 1,100 psi rupture discs and a target distance of 6 cm. Bombarded leaf samples were held in the dark at 25℃ for 2 days, cut into 1-cm
2 pieces, and placed adaxial side down onto regeneration MS medium containing 500 μg/mL spectinomycin dihydrochloride. Each bombardment was performed on approximately 20 explants. Only cells containing the plastid transformation vector with the spectinomycin resistance gene remained green and formed shoots, while nontransformed tissues lacking the resistance gene exhibited chlorosis and bleaching. The homologous integration of GFP-encoding genes into regenerated shoots was verified by PCR amplification using specific sets of primers (
Table 1). The shoots were subjected to an additional regeneration cycle on the same selective medium. Regenerated shoots were rooted in half-strength MS medium containing 500 μg/mL spectinomycin. Rooted plants were propagated in Jiffy peat pots before being transferred to the greenhouse for seed production (
Fig. 1,
Kumar & Daniell 2004,
Ruhlman et al. 2010). T
0 seeds from transplastomic plants were selected based on spectinomycin resistance and grown to produce T
1 seeds, which were subsequently used to analyze recombinant protein expression.
PCR was performed to confirm the presence of the target gene using gene-specific primers. Amplification was carried out in a 20-μL reaction containing 50 ng of genomic DNA using AccuPower PCR PreMix (Bioneer, Daejeon, South Korea), with an initial denaturation at 98℃ for 5 min, followed by 30 cycles of 98℃ for 30 s, 55℃ for 30 s, and 72℃ for 2 min and 30 s, with a final extension at 72℃ for 5 min. For cDNA synthesis, total RNA was extracted from tobacco leaves using the XENOPURE Total RNA Purification Kit (Cell To Bio, Seoungnam, Republic of Korea). For each construct, RNA was isolated from four independent T
1 transgenic plants, which were treated as biological replicates. First-strand cDNA was synthesized from 5 μg of total RNA using oligo (dT) primers and the RNA-to-cDNA EcoDry Premix (Takara, Shiga, Japan). Quantitative real-time PCR (qRT-PCR) was performed with 100 ng of cDNA as template in a 20 μL reaction using the AccuPower 2×GreenStar™ qPCR Master Mix (Bioneer, Daejeon, Republic of Korea). The thermal cycling protocol consisted of an initial denaturation at 95℃ for 10 min, followed by 40 cycles of 95℃ for 20 s, 58℃ for 20 s, and 72℃ for 25 s. Fluorescence was recorded at the end of each cycle. Each biological replicate was analyzed with three technical replicates. Relative gene expression levels were calculated using the 2
-ΔCt method (
Schmittgen & Livak 2008), with actin (UniProt accession no. Q05214) as the internal reference.
Leaves harvested from greenhouse-grown plants were frozen and ground into a fine powder with a mortar and pestle. TSP was extracted from 100 mg of this powder in 300 μL of plant extraction buffer, consisting of phosphate-buffered saline (PBS, 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, pH 7.4, supplemented with Roche EDTA-free complete protease inhibitor cocktail) for 1 h at 4℃. After extraction, TSP was quantified using the bicinchoninic acid assay (Thermo Fisher Scientific, Waltham, MA, USA, 23225) and run on Mini-PROTEAN TGX Precast gels (Bio-Rad, Hercules, CA, USA). Immunoblot analysis was performed using rabbit anti-GFP antibody (1:1,000, Abcam, Cambridge, UK, ab290) and goat anti-rabbit IgG-HRP secondary antibody (1:5,000, GenDEPOT, Katy, TX, USA, SA002-500). Recombinant GFP was used as a standard (ABclonal, Woburn, MA, USA, RTP0003). The relative intensities of the immunoblot bands were quantified using ImageJ software (NIH, Bethesda, MD, USA). Each band was selected using the rectangular selection tool, and the intensity profile was used to measure the area (signal intensity) of each band. The measured values were normalized to the GFP loading standard.
TSP samples from the four lines corresponding to each promoter were diluted and used for indirect enzyme-linked immunosorbent assay (ELISA). Recombinant GFP standards, samples, and blanks were coated onto wells in immunoplates (Thermo Fisher Scientific, Waltham, MA, USA) in serial dilutions with coating buffer (0.1 M NaHCO3, 0.03 M Na2CO3, pH 9.5). The immunoplate was then blocked with 1% bovine serum albumin in 1×PBS. For GFP antibody binding, the anti-GFP antibody (1:5,000, Abcam, Cambridge, UK, ab290) was used as the primary antibody, and goat anti-rabbit IgG-HRP secondary antibody (1:10,000, GenDEPOT, Katy, TX, USA, SA002-500) was used as the secondary antibody. To detect the signal, 100 μL of TMB (3, 3′, 5, 5′-tetramethylbenzidine) containing H2O2 solution was added, and the immunoplates were incubated for 20 min at room temperature. Color development was determined at a wavelength of 450 nm after adding a stopping solution (50 μL, 2 M H3PO4). The GFP level for each sample was calculated using a standard curve based on the recombinant GFP standards.
Results and Discussion
Construction of Chloroplast Expression Vectors
To compare the relative recombinant protein expression activity of the three different promoters, we constructed three plasmid vectors by cloning expression cassettes between the flanking sequences corresponding to the
trnI and
trnA genes of the tobacco chloroplast genome (
Fig. 2). The expression cassettes contained either the
rrn,
rbcL, or
psbA promoter, along with the coding sequence of GFP as a marker and the
psbA 3′ UTR to provide transcript stability.
aadA, which confers spectinomycin resistance, was used as the selectable marker.
Transplastomic tobacco lines were generated via biolistic transformation using the three promoter-specific vectors. After selection on spectinomycin-containing medium, four independent lines were obtained for each of the three promoter constructs. The transformation efficiency of the construct containing the
rrn promoter was 0.75%, whereas the efficiencies of the
rbcL and
psbA constructs were 0.68% and 0.63%, respectively (
Table 2). Although the
rrn construct showed a slightly higher point estimate, statistical tests revealed no significant differences among the three promoters (χ
2=0.15,
p=0.93). Thus, the apparent variation should be interpreted as an observational trend rather than a biologically meaningful difference. The average frequency for the
rrn construct corresponded to approximately 0.15 transplastomic events per bombardment. PCR analysis confirmed the site-specific integration of the transgene into the chloroplast genome in all selected lines (
Figs. 3A,
3B,
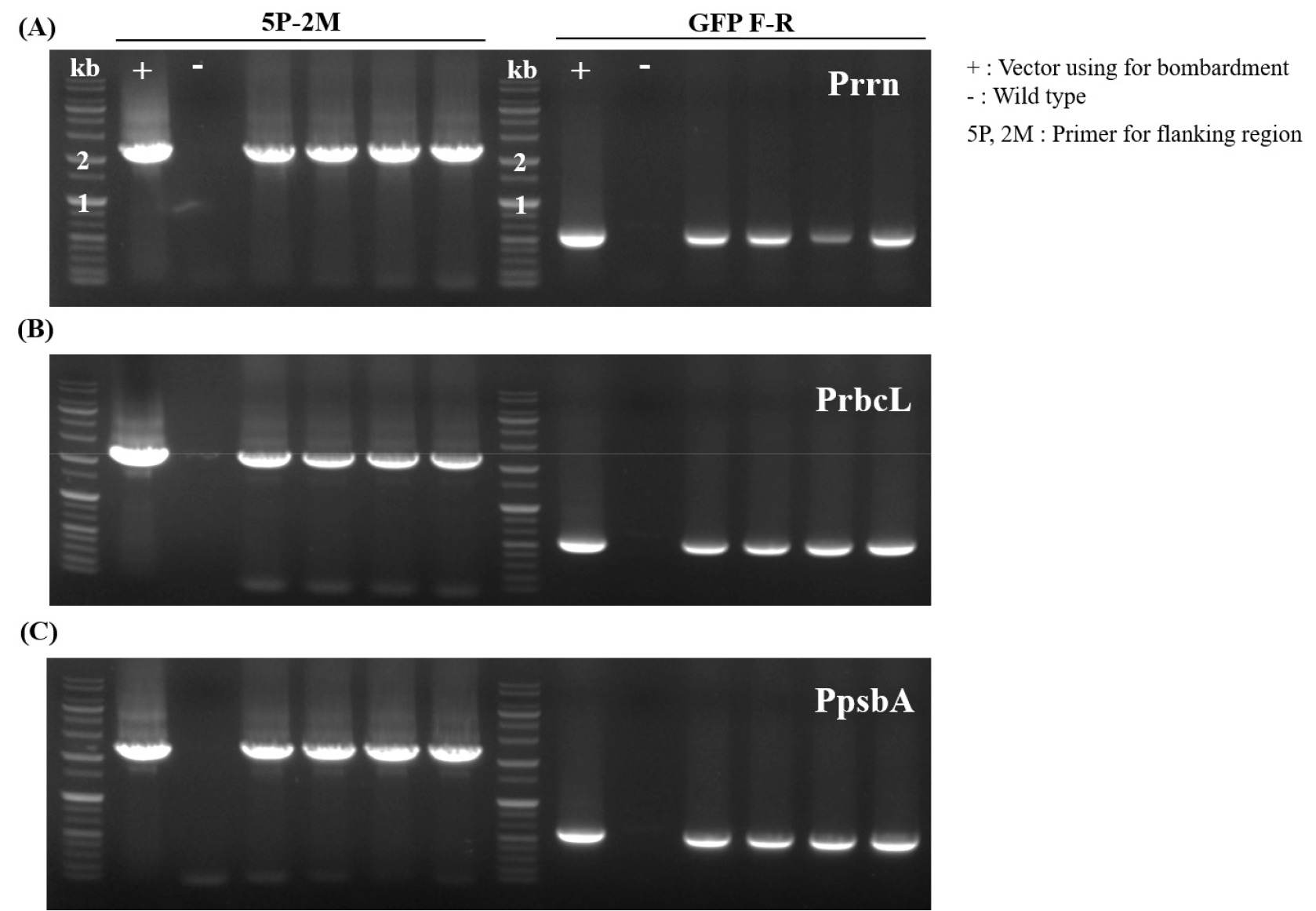
3C). To confirm site-specific integration of the transgene, PCR was performed using one primer annealing to the region outside the left (or right) flanking sequence in the chloroplast genome and another primer within the transgene cassette. Only transplastomic plants with the correct insertion produced the expected PCR product and wild-type plants yielded no product.
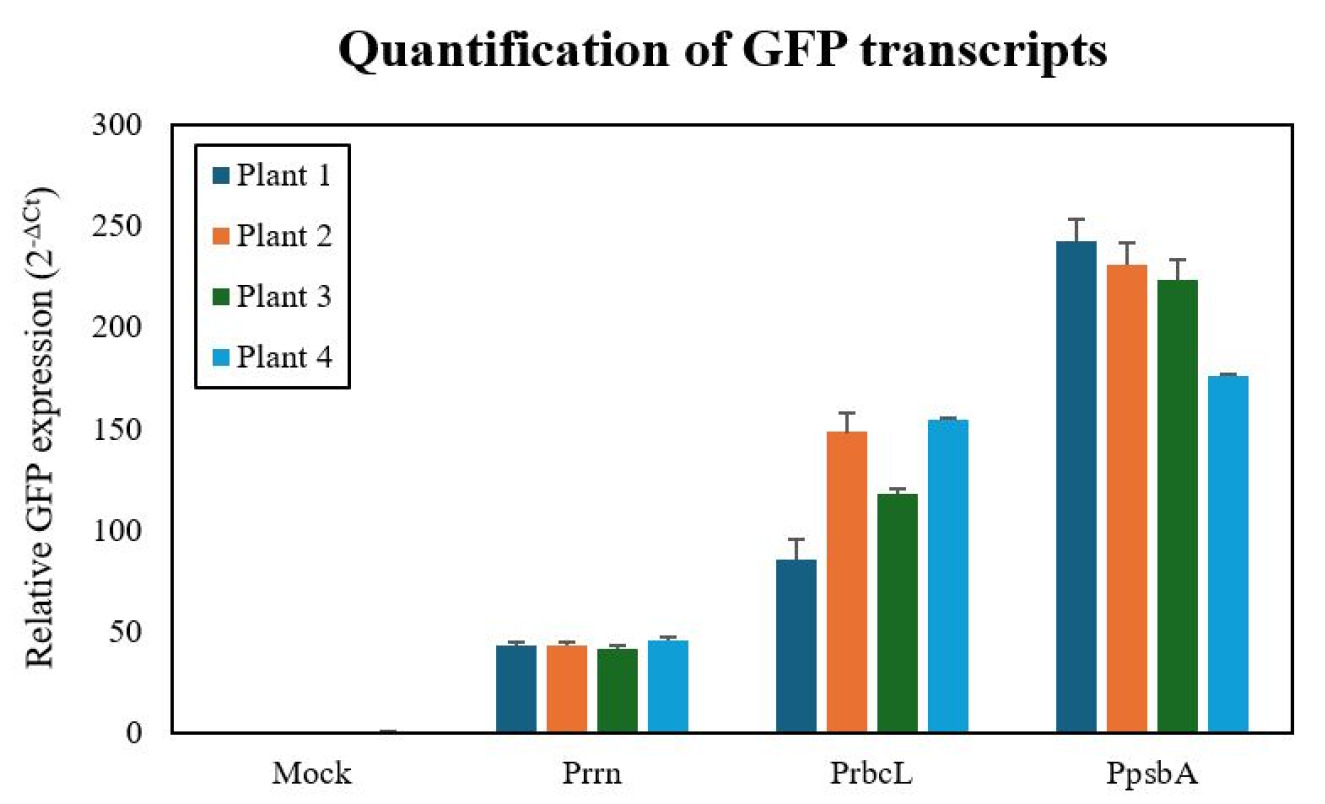
To compare the transcriptional activity of the three promoters, we quantified GFP mRNA levels by qRT-PCR using actin as a reference. The
psbA promoter exhibited the highest transcript levels, followed by
rbcL and
rrn, with mean relative expression values of 218.21±19.64, 126.60±8.78, and 43.27±1.57, respectively (
Fig. 4). One-way ANOVA confirmed that these differences were highly significant (F (2,6)=130.7,
p<0.000,
Armstrong et al. 2000). These results demonstrate that promoter choice strongly influences chloroplast transgene expression, with
psbA consistently driving the highest transcriptional activity under the conditions tested.
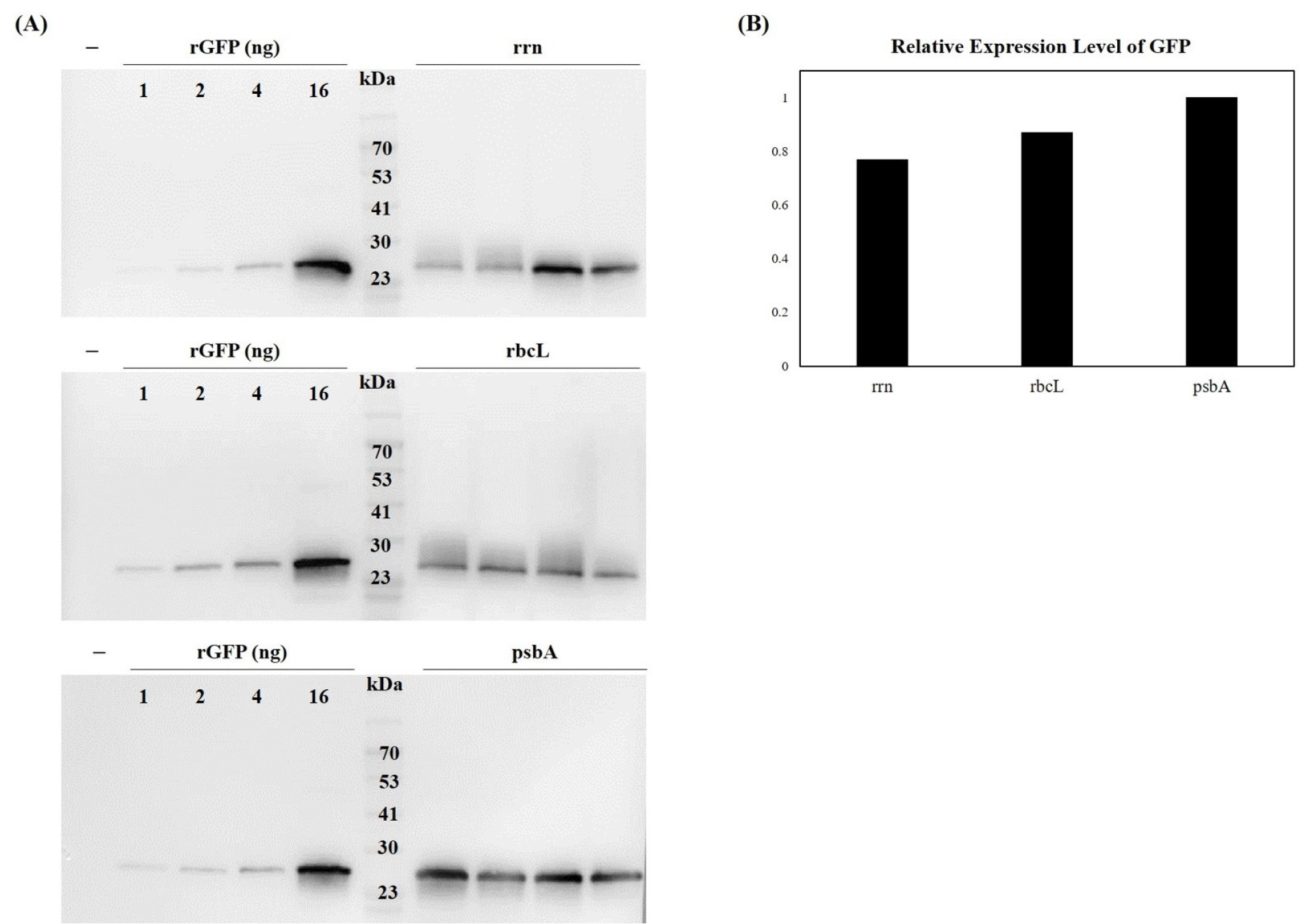
To correlate transcriptional activity with protein accumulation, we quantified GFP levels using both immunoblot and ELISA. Immunoblot analysis showed that the
psbA promoter produced the highest GFP signal, with weaker signals observed for the
rrn and
rbcL promoters (
Fig. 5). For quantitative analysis, recombinant GFP (rGFP) was used as a standard. Band intensities were measured using ImageJ software. Following normalization to the rGFP standard and then to the levels in
psbA, the relative intensity values for GFP were 1.00 (
psbA), 0.87 (
rbcL), and 0.77 (
rrn). These results indicated that the
psbA promoter drove the highest level of GFP protein accumulation, whereas the
rrn and
rbcL promoters resulted in approximately 13% and 23% lower expression, respectively.
To address the substantial variability observed among individual lines in both mRNA and immunoblot analyses, we performed ELISA measurements, which provided a representative assessment of GFP expression for each promoter group. Absolute GFP concentrations were determined independently from four individual transplastomic lines per construct and are presented as mean±standard deviation. The
psbA promoter group accumulated 616.2±28.7 ng GFP/g LFW, whereas the
rrn and
rbcL promoter groups accumulated 338.9±100.2 and 510.3±32.4 ng GFP/g LFW, respectively (
Fig. 6). One-way ANOVA revealed a significant difference among the three promoters (F (2,9)=19.17,
p=0.000,
Armstrong et al. 2000). On average, GFP expression driven by the
rrn and
rbcL promoters corresponded to approximately 55% and 83% of the
psbA promoter level, respectively. These replicate-based findings are in agreement with the immunoblot analysis, confirming that the
psbA promoter yielded the highest GFP protein levels. Taken together, both mRNA and protein analyses consistently demonstrated that the
psbA promoter conferred the most efficient expression of recombinant GFP in chloroplasts.
In this study, we conducted a systematic comparative evaluation of three chloroplast promoters-
psbA,
rbcL, and
rrn-to assess their efficacy in driving transgene expression in
N. tabacum. Using GFP as a reporter gene, we quantified expression levels at both transcriptional and translational levels through qRT-PCR, Western blotting, and ELISA. Among the three, the
psbA promoter consistently exhibited the highest expression levels, followed by
rbcL and
rrn. qRT-PCR analysis revealed that the
psbA promoter drove mRNA expression approximately fivefold and twofold higher than
rrn and
rbcL, respectively. This trend was mirrored by Western blot and ELISA results, indicating that transcriptional strength is a major determinant of recombinant protein yield in chloroplast systems. The
psbA promoter’s effectiveness stems from its role in regulating the D1 protein of Photosystem II, making it particularly efficient in green, photosynthetically active tissues (
Mulo et al. 2009). The
rbcL promoter showed moderate transcript levels but resulted in relatively high protein accumulation, potentially due to enhanced mRNA stability or translational efficiency. In contrast, the
rrn promoter showed the lowest expression at both transcript and protein levels, possibly due to suboptimal recognition by the plastid transcriptional machinery or a lack of necessary regulatory elements (
Suzuki et al. 2003). Despite being derived from the ribosomal RNA operon and known as one of the strongest chloroplast promoters in some contexts, its performance in this study was unexpectedly modest.
The transformation efficiency observed in this study was notably lower than previously reported values in the literature. While the
rrn promoter construct achieved the highest transformation efficiency at 0.75%, this figure is substantially below the up to 14 transplastomic lines were obtained per bombarded leaf for tobacco chloroplast transformation (
Daniell et al. 2001). Two factors may have contributed to the reduced transformation efficiency observed in this study. First, optimization of gold particle size and bombardment pressure parameters may have been insufficient, as previous studies have shown that 0.4 μm particles can increase transformation efficiency 3-4 folds compared to larger particles (
Langbecker et al. 2004), Second, the length and homology of flanking sequences used for homologous recombination may have been suboptimal (
Bock 2015). Future studies should optimize gold particle size (0.4-0.6 μm) and bombardment pressure (1,100-1,350 psi) to achieve standard transformation efficiencies. Additionally, optimizing the length and homology of flanking sequences could significantly improve transformation success rates.
The GFP expression levels obtained in this study (616.2±28.7 LFW for
psbA promoter) are considerably lower than those reported for other recombinant proteins produced through chloroplast transformation. Typically, chloroplast-expressed cellulolytic enzymes, xylanases attained 1.3 mg/g LFW (
Kolotilin et al. 2013) and human EGF achieve 1.57 ug/g LFW (
Wang et al. 2023). The low GFP expression levels can be attributed to two factors specific to fluorescent proteins. First, protein folding and stability issues may affect GFP expression in the chloroplast environment (
Foyer & Hanke 2022,
Llamas & Pulido 2022). GFP requires formation of a β-barrel structure to exhibit fluorescence, and this structural formation may be inefficient in the oxidative environment of chloroplasts (
Reid & Flynn 1997). The presence of cysteine residues in GFP can be particularly problematic in the chloroplast’s oxidative environment, potentially interfering with proper protein folding (
Aronson et al. 2011). Second, protein degradation pathways may specifically target GFP in chloroplasts (
Apel et al. 2010). Chloroplast-localized proteins can be subject to N-end rule degradation pathways, and the N-terminal amino acid composition can significantly affect protein half-life in this compartment (
Bouchnak & van Wijk 2019).
This study has several limitations that should be acknowledged. First, all experiments were conducted exclusively in
N. tabacum, and promoter performance in other crop species remains unverified. Second, only GFP was tested as a reporter, and performance may vary significantly for structurally or functionally different protein. Third, homoplasmy achievement and growth characteristics under selection pressure were not evaluated. Fourth, as discussed above, transformation efficiency was substantially lower than literature-reported values, indicating a need for protocol optimization (
Daniell et al. 2001). Fifth, the low GFP expression levels may be attributed more to GFP-specific characteristics than to promoter efficiency limitations. Future research should include promoter performance comparisons across various plant species, environmental stress condition expression pattern analysis, and applicability verification for diverse target proteins. Particularly, studies using codon-optimized GFP or comparative analyses with other reporter proteins would provide more accurate promoter efficiency evaluations. Additionally, synthetic or hybrid promoter development combining favorable characteristics such as strong basal activity, inducibility, and transcript stability represents an important research direction (
Inckemann et al. 2024).
The promoter hierarchy confirmed in this study (
psbA>
rbcL>
rrn) provides useful guidelines for expression system design across various application fields. The
psbA promoter is suitable for applications requiring high expression levels, such as biopharmaceuticals and industrial enzyme, potentially contributing to commercial cost-efficiency improvements (
Rasala et al. 2011). Plant-based molecular farming can achieve annual production scales of hundreds of kilograms to several tons of recombinant proteins, providing economic advantages over traditional microbial or animal cell culture systems (
Eidenberger et al. 2023,
Schillberg et al. 2019). The
rbcL promoter would be useful for moderate expression levels or situations where overaccumulation is detrimental (
Shiina et al. 1998). The
psbA promoter’s light and environmental stimulus responsiveness can be leveraged for spatiotemporal or conditional expression, helping minimize metabolic burden during early growth stages or in non-photosynthetic tissues. Ultimately, realizing the industrial potential of chloroplast biotechnology will require integration of promoter optimization with stable transgene insertion, marker-free transformation, and scalable cultivation strategies.
Conclusion
This study quantitatively demonstrated the importance of promoter selection for recombinant protein production in chloroplast transformation. The superior performance of the psbA promoter provides a foundation for developing high-efficiency chloroplast expression systems that will advance plant-based molecular farming. Transformation efficiency results fell short of other reported values, indicating an area in need of improvement. Additionally, the low expression levels of GFP suggested a need to optimize codon use, while addressing protein stability issues and inherent characteristics of fluorescent proteins in future research. Overall, the research results and limitations presented here provide a foundation for rational design of scalable, efficient, and cost-effective chloroplast-based biofactories.
Acknowledgments
We gratefully acknowledge Dr. Seung-Bum Lee for generously providing the pBL vector used in our experiments. This research was supported by a research grant from the Rural Development Administration project (Project No. PJ01724502, JL), Republic of Korea.
Fig. 1Photographic overview of the chloroplast transformation process in N. tabacum using particle bombardment. (A) Leaf pieces transferred to selective medium containing 500 μg/mL spectinomycin (B) Leaf disks showing bleaching of non-transformed tissue while resistant sectors remain green (C), (D) regenerating shoots on selective medium (E) Acclimatization process following successful root induction, with rooted transplastomic plantlets being prepared for greenhouse transfer (F) Transplastomic plants immediately after transfer to isolated GMO greenhouse conditions in individual pots for containment and further evaluation (G) Well-established transplastomic plants thriving in greenhouse pots, demonstrating normal vegetative growth and development. (H) Mature transplastomic plants that have progressed to flowering stage, showing no adverse phenotypic effects and normal reproductive development, confirming successful chloroplast transformation.

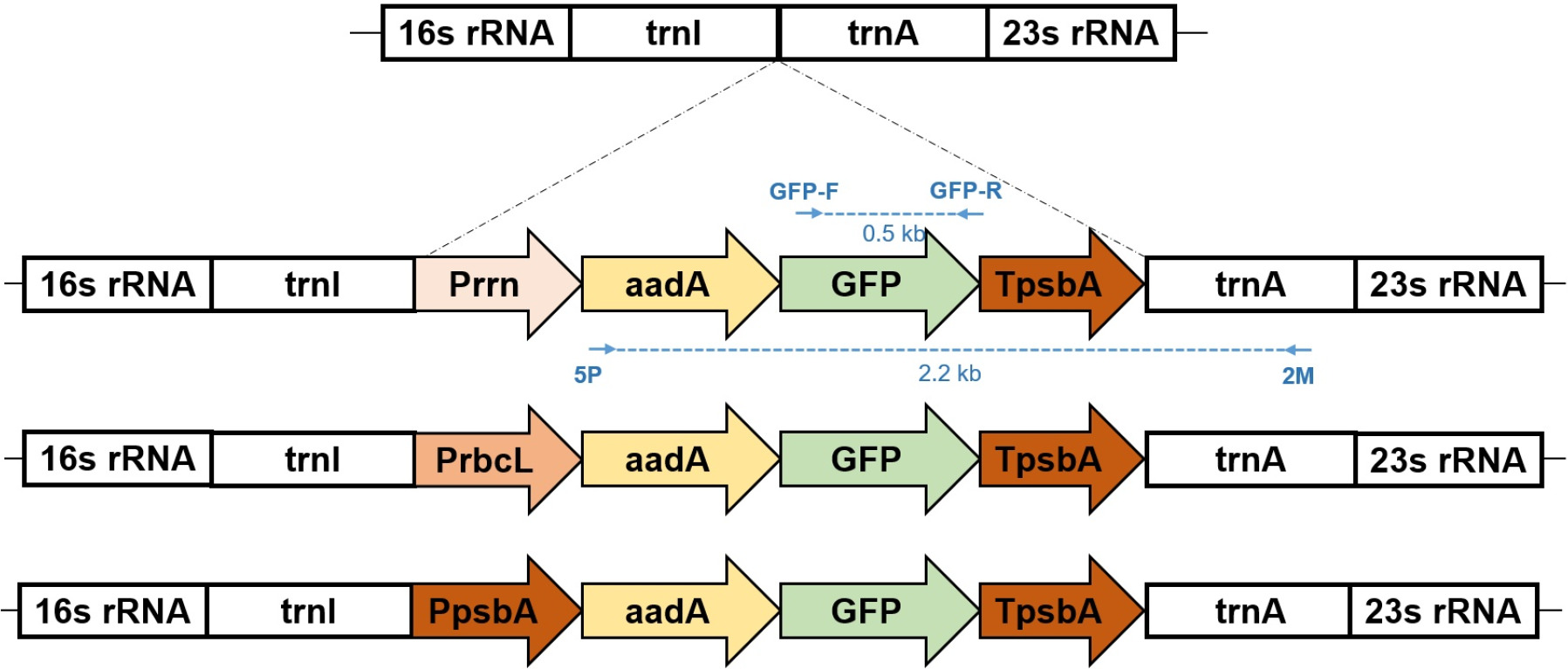
Fig. 2Schematic representation of three chloroplast transformation vectors for GFP expression. The vectors contain GFP coding sequences driven by three different promoters: (A) rrn, (B) rbcL and (C) psbA. Each construct includes flanking sequences (trnI and trnA genes) for homologous recombination into the tobacco chloroplast genome, the respective promoter region, GFP coding sequence, psbA 3’ UTR for transcript stability, and aadA selectable marker gene conferring spectinomycin resistance. The arrows indicate the direction of transcription. PCR primer binding sites are indicated: 5P-2M primer (forward) and GFP-F/R primer pair for transgene verification. The expected PCR products generated by these primer sets are shown with skyblue dotted lines, enabling confirmation of successful integration and proper orientation of the GFP transgene within the chloroplast genome.
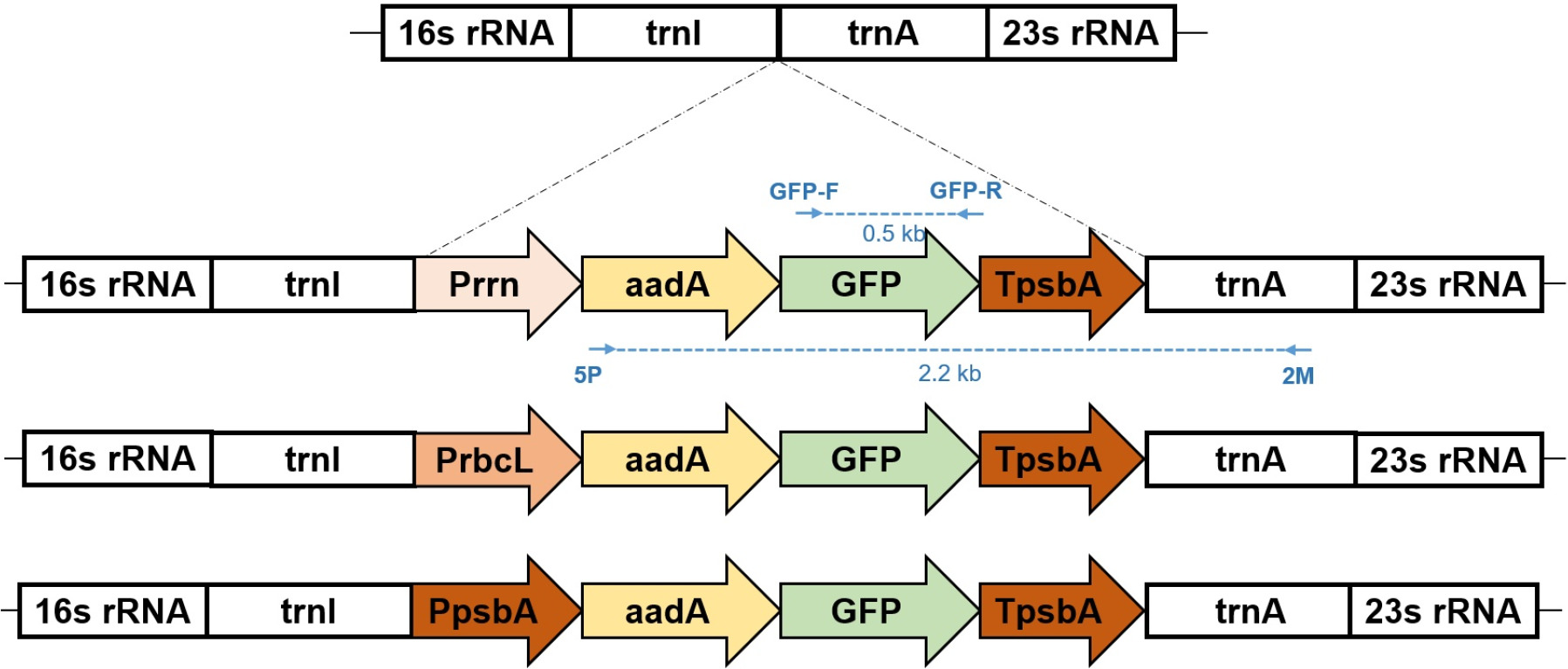
Fig. 3PCR analysis of transplastomic N. tabacum lines expressing GFP under different promoters. Genomic DNA from four independent lines each was analyzed by PCR using specific primer sets to confirm transgene integration into the chloroplast genome. (A) Lines transformed with rrn promoter construct, (B) lines with rbcL promoter construct, and (C) lines with psbA promoter construct. All selected lines showed positive PCR amplification, confirming successful integration of GFP transgenes into the chloroplast genome.
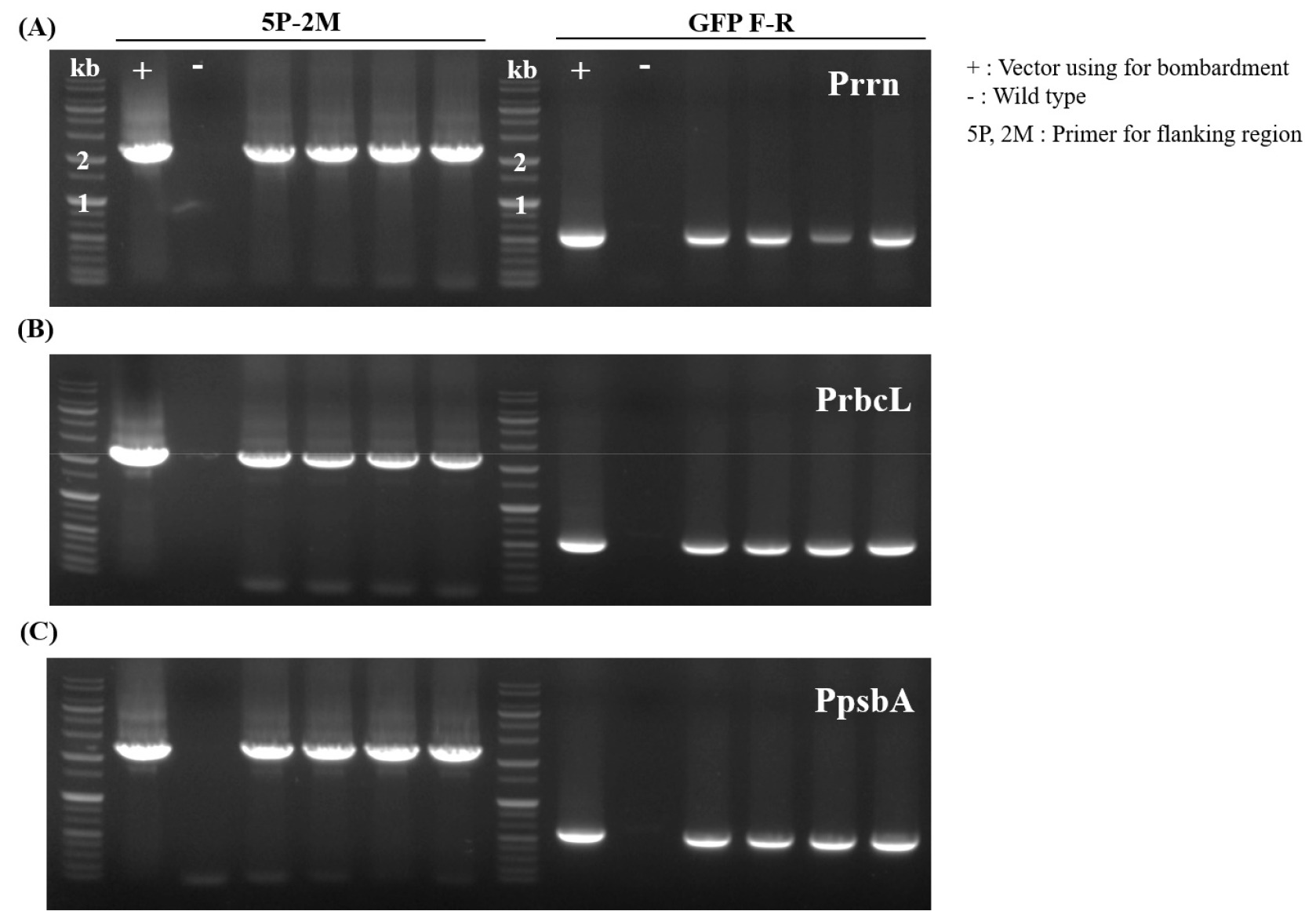
Fig. 4Quantitative analysis of GFP mRNA expression in transplastomic N. tabacum leaves. Real-time PCR analysis comparing GFP transcript levels among three promoter groups. Each bar represents mean relative expression±standard deviation (n=3) normalized to actin reference gene. The psbA promoter group showed significantly higher GFP mRNA levels (218.21±19.64) compared to rbcL (126.60±8.78) and rrn (43.27±1.57) promoter groups. One-way ANOVA revealed statistically significant differences among groups (F (2,6)=130.7, p<0.000).
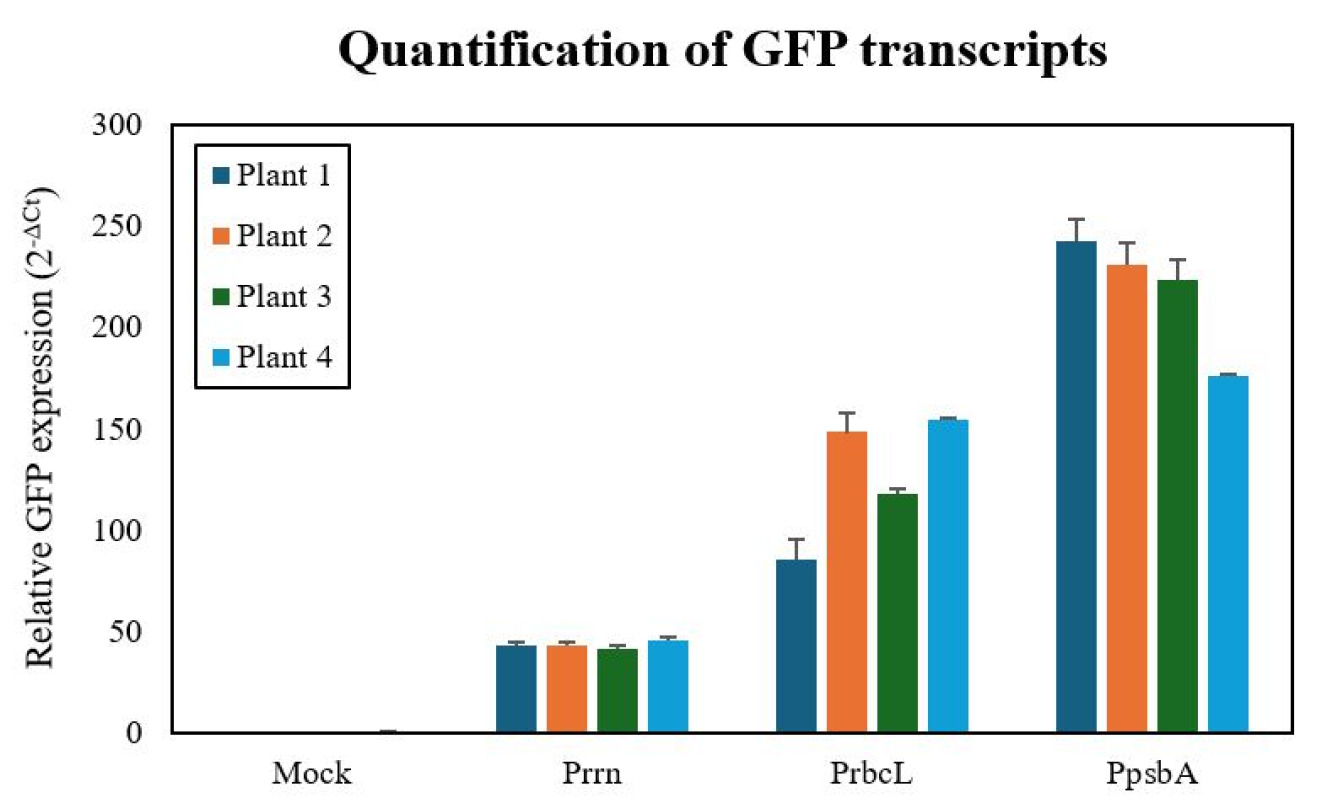
Fig. 5Western blot analysis of GFP expression under three different chloroplast promoters. (A) Total soluble proteins were extracted from transplastomic tobacco leaves and analyzed by SDS-PAGE followed by immunoblotting with anti-GFP antibody (1:1,000 dilution). Recombinant GFP (rGFP) was used as positive control and molecular weight standard. (B) Band intensities were quantified using ImageJ software and normalized to rGFP standard, showing relative expression levels of 1.00 (psbA), 0.89 (rbcL), and 0.75 (rrn). The psbA promoter drove the highest GFP protein accumulation levels.
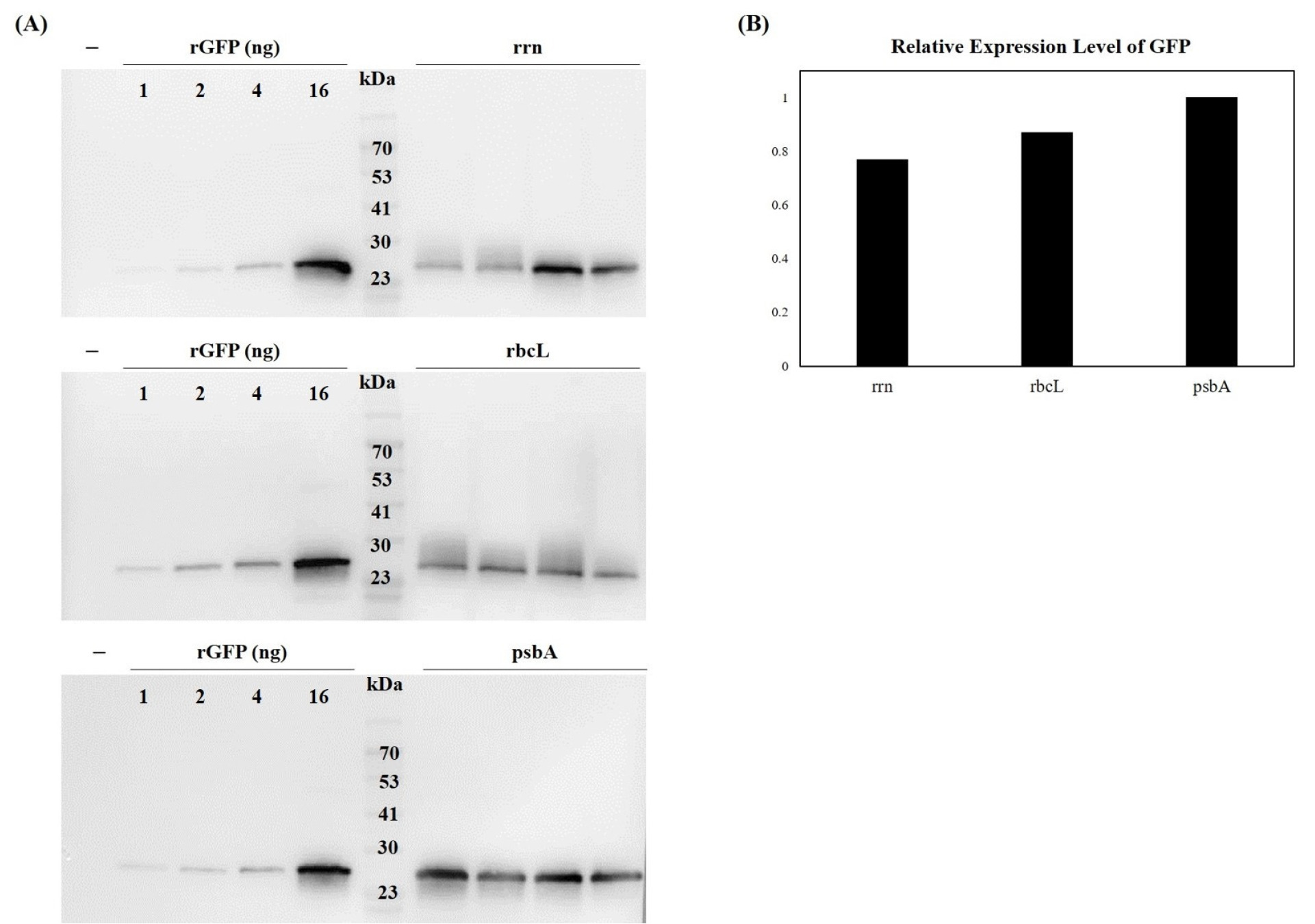
Fig. 6Quantitative analysis of GFP levels using ELISA. Absolute GFP concentrations were determined by ELISA from four independent transplastomic lines for each promoter construct (n=4 biological replicates). Data are presented as mean±standard deviation. The psbA promoter group accumulated 616.2±28.7 ng GFP/g LFW, whereas the rrn and rbcL promoter groups accumulated 338.9±100.2 and 510.3±32.4 ng GFP/g LFW, respectively. On average, GFP expression driven by the rrn and rbcL promoters corresponded to approximately 55% and 83% of the psbA promoter level. These results demonstrate that the psbA promoter confers the highest GFP protein accumulation in chloroplasts, consistent with immunoblot and transcript analyses.
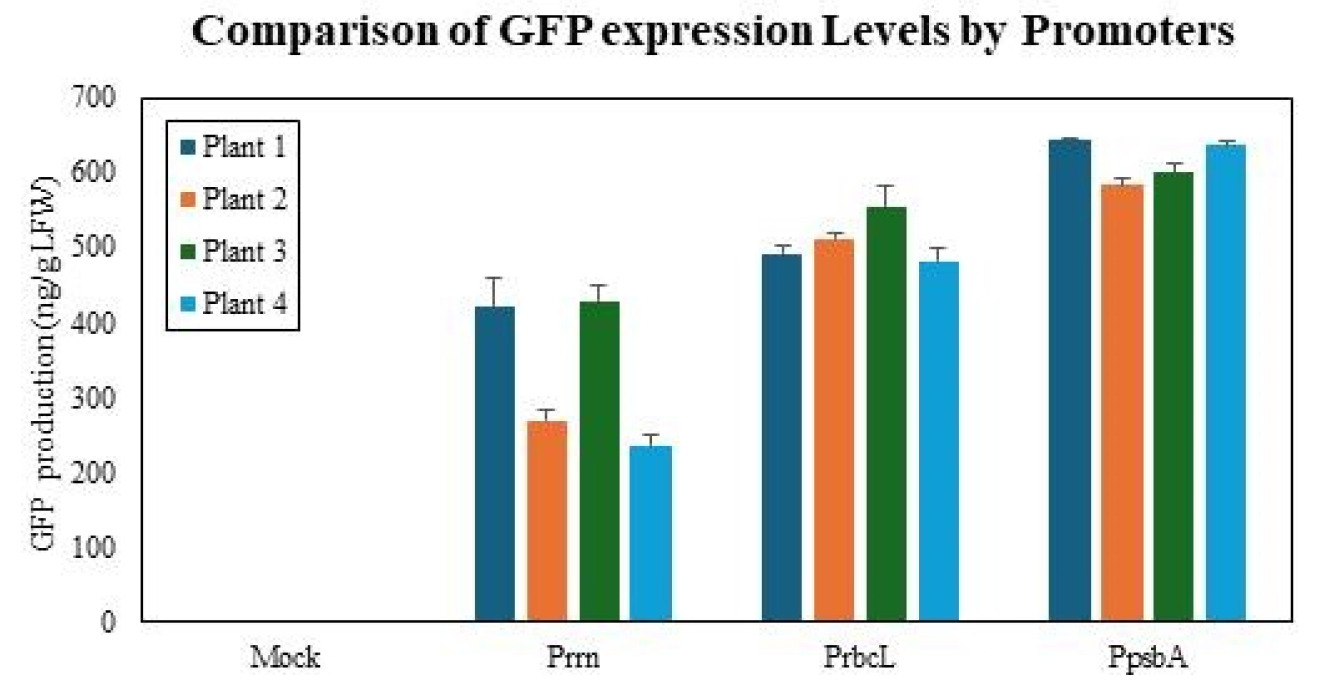
Table 1Primer sequences (5'-3') used for PCR analysis of GFP insertion in the chloroplast genome.
Table 1
|
Primer |
Sequence (5'-3') |
|
5P |
ctgtagaagtcaccattgttgtgc |
|
2M |
tgactgcccaacctgagagc |
|
GFP-F |
agtggagagggtgaaggtga |
|
GFP-R |
aaagggcagattgtgtggac |
Table 2Transformation efficiency of chloroplasts from N.tabacum with three different vectors.
Table 2
|
Construct |
Leaf disc bombarded |
Shoots regenerated |
Positive plants |
Efficiency (%)z
|
The pBL
Prrn-GFP-TpsbA |
1876 |
28 |
14 |
0.75 |
The pBL
PrbcL-GFP-TpsbA |
1169 |
24 |
8 |
0.68 |
The pBL
PpsbA-GFP-TpsbA |
1268 |
21 |
8 |
0.63 |
References
- 1. Allison LA, Simon LD, Maliga P. 1993. Deletion of rpoB reveals a second distinct transcription system in plastids of higher plants. EMBO J 12: 2807-2814.
- 2. Apel W, Schulze WX, Bock R. 2010. Identification of protein stability determinants in chloroplasts. Plant J 63: 636-650.
- 3. Armstrong RA, Slade SV, Eperjesi F. 2000. An introduction to analysis of variance (ANOVA) with special reference to data from clinical experiments in optometry. Ophthalmic Physiol Opt 20: 235-241.
- 4. Aronson DE, Costantini LM, Snapp EL. 2011. Superfolder GFP is fluorescent in oxidizing environments when targeted via the Sec translocon. Traffic 12: 543-548.
- 5. Bock R, Chan HT, Daniell H. 2018. Chloroplast biotechnology: Present and future. Science 360: 392-393.
- 6. Bock R. 2015. Engineering plastid genomes: Methods, tools, and applications in basic research and biotechnology. Annu Rev Plant Biol 66: 211-241.
- 7. Bouchnak I, van Wijk KJ. 2019. N-Degron pathways in plastids. Trends Plant Sci 24: 917-926.
- 8. Castiglia D, Sannino L, Marcolongo L, Ionata E, Tamburino R, De Stradis A, Cobucci-Ponzano B, Moracci M, La Cara F, Scotti N. 2016. High-level expression of thermostable cellulolytic enzymes in tobacco transplastomic plants and their use in hydrolysis of an industrially pretreated Arundo donax L. biomass. Biotechnol Biofuels 9: 154
- 9. Cheon KS, Jeong IS, Kim KH, Lee MH, Lee TH, Lee JH, Yoon UH, Chandra R, Lee YJ, Chandra R, Kim TH. 2018. Comparative SNP analysis of chloroplast genomes and 45S nrDNAs reveals genetic diversity of Perilla species. Plant Breed Biotech 6: 125-139.
- 10. Daniell H, Lee SB, Panchal T, Wiebe PO. 2001. Expression of the native cholera toxin B subunit gene and assembly as functional oligomers in transgenic tobacco chloroplasts. J Mol Biol 311: 1001-1009.
- 11. Daniell H, Lin CS, Yu M, Chang WJ. 2016. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol 17: 134
- 12. Daniell H, Singh ND, Mason H, Chan HT. 2020. Plant-made oral biological therapeutics: An attractive alternative to injectable biologics. Plant Biotechnol J 18: 1627-1642.
- 13. Daniell H, Streatfield SJ, Wycoff K. 2001. Medical molecular farming: Production of antibodies, biopharmaceuticals and edible vaccines in plants. Trends Plant Sci 6: 219-226.
- 14. Eidenberger L, Kogelmann B, Steinkellner H. 2023. Plant-based biopharmaceutical engineering. Nat Rev Bioeng 1: 426-439.
- 15. Feng Z, Li X, Fan B, Zhu C, Chen Z. 2022. Maximizing the production of recombinant proteins in plants: From transcription to protein stability. Int J Mol Sci 23: 13516
- 16. Foyer CH, Hanke G. 2022. ROS production and signalling in chloroplasts: Cornerstones and evolving concepts. Plant J 111: 642-661.
- 17. Giddings G. 2001. Transgenic plants as protein factories. Curr Opin Biotechnol 12: 450-454.
- 18. Inckemann R, Chotel T, Brinkmann CK, Burgis M, Andreas L, Baumann J, Sharma P, Klose M, Barret J, Ries F, Paczia N, Glatter T, Willmund F, Mackinder LCM, Erb TJ. 2024. Advancing chloroplast synthetic biology through high-throughput plastome engineering of Chlamydomonas reinhardtii. bioRxiv
- 19. Kolotilin I, Kaldis A, Pereira EO, Laberge S, Menassa R, Matthew M. 2013. Optimization of transplastomic production of hemicellulases in tobacco: Effects of expression cassette configuration and tobacco cultivar used as production platform on recombinant protein yields. Biotechnol Biofuels 6: 65
- 20. Kumar S, Daniell H. 2004. Engineering the chloroplast genome for hyperexpression of human therapeutic proteins and vaccine antigens. Methods Mol Biol 267: 365-382.
- 21. Langbecker CL, Ye GN, Broyles DL, Duggan LL, Xu CW, Hajdukiewicz PTJ, Armstrong CL, Staub JM. 2004. High-frequency transformation of undeveloped plastids in tobacco suspension cells. Plant Physiol 135: 39-46.
- 22. Lee J, Lee SK, Park JS, Lee KR. 2023. Plant-made pharmaceuticals: Exploring studies for the production of recombinant protein in plants and assessing challenges ahead. Plant Biotechnol Rep 17: 53-65.
- 23. Legen J, Duhnen S, Gauert A, Gotz M, Schmitz-Linneweber C. 2023. A CRR2-dependent sRNA sequence supports papillomavirus vaccine expression in tobacco chloroplasts. Metabolites 13: 315
- 24. Llamas E, Pulido P. 2022. A proteostasis network safeguards the chloroplast proteome. Essays Biochem 66: 219-228.
- 25. Maliga P. 2004. Plastid transformation in higher plants. Annu Rev Plant Biol 55: 289-313.
- 26. Mulo P, Sicora C, Aro EM. 2009. Cyanobacterial psbA gene family: Optimization of oxygenic photosynthesis. Cell Mol Life Sci 66: 3697-3710.
- 27. Oey M, Lohse M, Kreikemeyer B, Bock R. 2009. Exhaustion of the chloroplast protein synthesis capacity by massive expression of a highly stable protein antibiotic. Plant J 57: 436-445.
- 28.
- 29. Rasala BA, Muto M, Sullivan J, Mayfield SP. 2011. Improved heterologous protein expression in the chloroplast of Chlamydomonas reinhardtii through promoter and 5′ untranslated region optimization. Plant Biotechnol J 9: 674-683.
- 30. Reid BG, Flynn GC. 1997. Chromophore formation in green fluorescent protein. Biochemistry 36: 6786-6791.
- 31. Ruhlman T, Verma D, Samson N, Daniell H. 2010. The role of heterologous chloroplast sequence elements in transgene integration and expression. Plant Physiol 152: 2088-2104.
- 32. Sambrook J, Russell DW. 2001. Molecular cloning: A laboratory manual. 3rd ed. Cold Spring Harbor Laboratory Press.
- 33. Schillberg S, Raven N, Spiegel H, Rasche S, Buntru M. 2019. Critical analysis of the commercial potential of plants for the production of recombinant proteins. Front Plant Sci 10: 720
- 34. Schmittgen TD, Livak KJ. 2008. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc 3: 1101-1108.
- 35. Shiina T, Allison L, Maliga P. 1998. rbcL transcript levels in tobacco plastids are independent of light: Reduced dark transcription rate is compensated by increased mRNA stability. Plant Cell 10: 1713-1722.
- 36. Staub JM, Maliga P. 1993. Accumulation of D1 polypeptide in tobacco plastids is regulated via the untranslated region of the psbA mRNA. EMBO J 12: 601-606.
- 37. Staub JM, Maliga P. 1994. Translation of psbA mRNA is regulated by light via the 5′-untranslated region in tobacco plastids. Plant J 6: 547-553.
- 38. Su H, van Eerde A, Rimstad E, Bock R, Branza-Nichita N, Yakovlev IA, Clarke JL. 2023. Plant-made vaccines against viral diseases in humans and farm animals. Front Plant Sci 14: 1170815.
- 39. Suzuki JY, Sriraman P, Svab Z, Maliga P. 2003. Unique requirements for translation of the plastid rbcL mRNA. Plant J 34: 873-882.
- 40. Svab Z, Maliga P. 1993. High-frequency plastid transformation in tobacco by selection for a chimeric aadA gene. Proc Natl Acad Sci USA 90: 913-917.
- 41. Venkataraman S, Khan I, Habibi P, Le M, Lippert R, Hefferon K. 2023. Recent advances in expression and purification strategies for plant made vaccines. Front Plant Sci 14: 1273958.
- 42. Wang Y, Fan J, Wei Z, Xing S. 2023. Efficient expression of fusion human epidermal growth factor in tobacco chloroplasts. BMC Biotechnol 23: 1