적요
Suppression subtractive hybridization was carried out to identify resistant genes against to pear scab caused by Venturia nashicola using leaves harvested at 24 and 48 hours after inoculation into ‘93-3-98’ (highly resistant) and ‘Sweat Skin’ (highly susceptible). As a result of the ESTs analysis, nine and 14 unique genes were expressed on 24H (tester, mRNA at 24hr after inoculation of ‘93-3-98’; driver, one of ‘Sweet Skin’) and 48H (tester, mRNA at 48hr after inoculation of ‘93-3-98’; driver, one of ‘Sweet Skin’), respectively and genes related to defense or stress response were accounted for 40% (24H) and 42% (48H). Differential expressed genes classifying into PR protein family were pathogenesis-related protein 1a, major allergen Pyr c1 and allergen mal d 1 at 24H and major allergen Mal d 1.03B at 48H, respectively. Major allergen Pyr c1, F-Box/kelch-repeat protein, Flavoprotein wrbA, and hypothetical protein POPTRDRAFT_783792 are expected to closely connecting to scab resistance of pear following strongly expressed in highly resistant cv. ‘Bartlett’ and ‘93-3-98’ compared with moderately susceptible cv. ‘Gamcheonbae’, susceptible cv. ‘Wonhwang’, and highly susceptible cv. ‘Niitaka’ and ‘Sweat skin’.
서 언
일반적으로 식물체가 병원체에 노출되거나 세포가 손상되 면 특정 물질들이 용출되며 이를 감지하여 방어반응을 가지 게 된다. 이러한 반응은 systemic acquired resistance (SAR) 이라고 알려진 방어 메커니즘으로 병원균에 의해 손상된 식 물조직이 다른 조직으로 salicylic acid나 jasmonic acid, ethylene과 같은 물질을 보내어 식물이 전체적으로 방어반응을 유도하며, 감염된 조직으로부터 superoxide, peroxide, hydrogen peroxide와 같은 활성산소를 방출하여 병원균을 사멸시킨다 (
Sticher et al. 1997). 또한 감염된 부위의 세포들을 사멸시켜 병원균이 생장하지 못하게 하는 과민성반응과 같은 방법을 이용하기도 한다(
Hammond-Kosack & Jones 1996). 이러한 SAR 메커니즘에 따라 병원체에 공격받은 식물세포에서 β -1,3-glucanase (PR-1 family), endo-chitinase (PR-3 family), 그리고 thaumatin-like protein (PR-5 family)와 같은 pathogen related (PR) protein의 발현이 유도되어지며(
Paparu et al. 2007) 그 밖에 thionin, lipoxygenase, proteinase inhibitor, ACC oxidase, peroxidase, glutathione s-transferase와 같은 유전 자들의 발현이 증가됨이 보고되어 있다(
Hammond-Kosack & Jones 1996).
배 검은별무늬병 저항성에 관련된 유전자를 탐색하기 위한 방법으로 suppression subtractive hybridization (SSH)을 이 용하였다. 이는 대조구와 처리구의 전사체 사이에 차등 발현 되는 cDNA 단편을 PCR을 이용하여 선택적으로 증폭하는 기술로써 비특이적 증폭은 억제하고 특이적인 cDNA만을 효 과적으로 증폭시켜서 차등 발현 유전자를 선별하게 된다. 이 분석방법은 housekeeping 유전자의 증폭을 억제하면서 발현 되는 전사체 만을 선택적으로 증폭할 수 있으며 분리하고자 하는 유전자의 염기서열과 같은 정보 없이도 원하는 유전자 를 분리할 수 있는 장점이 있다. 또한 특수한 장비나 분석이 요구되지 않고 일반적인 분자생화적 기술 만으로도 분석이 가 능하기 때문에(
Moody 2001) 다양한 식물에서 병 저항성에 관 여하는 유전자를 분리하기 위하여 사용되고 있다(
Degenhardt et al. 2005,
Pan et al. 2005,
Xiong et al. 2001).
본 연구에서는 배 검은별무늬병에 고도 저항성으로
Rvn2 유전자 연관 DNA marker (
Cho et al. 2009) 개발에 저항성 친으로 사용한 ’93-3-98’(‘단배’ × ‘Bartlett’)에서 일정하게 높은 수준으로, 감수성 품종 ‘스위트스킨’에서는 낮은 수준으 로 전사되는 유전자들을 선발하고 염기서열 분석을 통하여 배 검은별무늬병 저항성에 관련된 유전자들을 탐색하였고 배 검은별무늬병 저항성에 차이를 보이는 품종들에서 선발 유전 자들의 발현을 분석하여 후보 유전자를 선발하였다.
재료 및 방법
식물 재료
V. nashicola에 고도 저항성 ‘93-3-98’과 고도 감수성 ‘스 위트스킨’을 시험재료로 사용하였다. 시험수는 원예작물을 위 한 상업적 배지(서울바이오, 순천, 한국)로 채워져 있는 7.6 L 플라스틱 포트에 재식된 P. pyrifolia 대목에 접목하여 병 처 리까지 주야온도가 30°C/12°C로 유지되는 유리온실에 보존 하였다. V. nashicola 접종 후 저항성과 감수성간의 차등발현 전사체를 SSH 방법으로 선발하기 위하여 전엽 20일 미만의 유엽을 사용하였다. 차등발현 유전자의 발현은 V. nashicola에 대하여 저항성 정도가 다른 ‘Bartlett’(고도 저항성, non-host), ‘93-3-98’(고도 저항성), ‘황실리’(저항성), ‘감천배’(중도 저 항성), ‘원황’(감수성), ‘신고’(고도 감수성), ‘스위트스킨’(고 도 감수성)간 비교하였다.
분생포자 인공접종 및 시료채취
전남 나주 배시험장의 검은별무늬병이 발병된 ‘신고’ 잎에 서 분생포자를 채취하였고 멸균수로 현탁하여 -80°C에 냉동 보관하였다가 이것을 약 20°C에서 녹인 후 0.1% sucrose와 Tween 80을 첨가하여 접종원으로 사용하였다. 접종원을 1 × 105 분생포자/mL 농도로 10개 내외의 잎이 완전히 전개된 1 년생 접목묘(6주이상/품종)의 수관 전체에 분무 접종하였으며 대조구 식물체는 0.1% sucrose와 0.005% Tween 80을 포함 하는 멸균수를 분무하였다. 접종은 모든 품종에서 동시에 처 리하였으며 접종 후 각 접목묘를 20°C, 상대습도 100% 습실 내에서 48시간을 유지한 후 50% 정도 차광된 실외에 두었다. 접종한 잎과 접종하지 않은 잎은 24시간 간격으로 0, 24, 48, 72, 96, 120시간 샘플을 채취하여 액체질소에 담근 후 -80°C의 냉동 고에 보관하였다(ilShinBioBase, Dongducheon, Kyeonggi-do, Korea)
RNA 분리 및 cDNA 합성
동결된 잎으로부터 RNA을 추출하기 위하여 변형된 cetyltrimethylammonium bromide (CTAB) 방법과 lithium chloride (LiCl) 침전방법을 사용하였다(
Iandolino et al. 2004). 잎은 액체질소에서 마쇄하여 -80°C에서 보관하였으며 RNA분리에 사용하였다. 마쇄된 조직은 CTAB buffer[2% CTAB, 2% polyvinlypyrrolidone, 100 mM Tris-HCl (pH 8.0), 25 mM EDTA, 2M NaCl]를 첨가하여 균질화 하였으며 65°C에서 30분 간 반응 후 순도를 높이기 위하여 두 번의 chloroform/isoamyl alcohol (IAA)(24:1)를 처리하여 상징액을 추출하였다. 3M LiCl를 첨가한 후 4°C에서 overnight처리하여 RNA를 농축 하였으며 RNA pellet은 sodium dodecyl sulfate–Tris-HCl– EDTA (SSTE) buffer [1M NaCl, 0.5% SDS, 10 mM Tris-HCl (pH 8.0), and 1mM EDTA (pH 8.0)]를 넣어 완전 히 용해하였다. Chloroform을 처리하여 상등액을 추출하였으 며 100% 에탄올 2 (v/v)을 첨가한 후 -20°C에서 30분간 반응 시켜 RNA를 농축하였다. Formaldehyde gel 전기영동을 이 용하여 분리된 RNA의 분해 정도를 확인하였으며 Agarose gel 전기영동을 통하여 DNA가 검출되지 않음을 확인하였다. 분리된 RNA는 spectrophotometer 260/280 nm에서 정량한 후 SuperScript III First-Strand Synthesis System for RT-PCR (Invitrogene, Waltham. Messachusetts, USA)을 사용하여 1
μg의 RNA로부터 cDNA를 합성하였다.
PCR select cDNA subtraction kit (Clontech, Mountain View, CA, USA)를 사용하여 라이브러리를 제작하였으며 SMART cDNA synthesis kit (Clontech)를 사용하여 total RNA 2
μg으로 부터 cDNA를 합성하였다. 저항성에서만 차등 발현되는 다양한 유전자를 탐색하기 위하여 분생포자의 발아 관과 균사의 신장 최성기로 병과 식물체와 상호작용이 활발 한 시기인(
Park et al. 2000), 24H(‘93-3-98’ 24시간 vs ‘스위 트스킨’ 24시간), 48H(‘93-3-98’ 48시간 vs ‘스위트스킨’ 48시 간)로 나누어 suppression subtractive hybridization (SSH)를 수행하였다. 저항성 품종에서 차등 발현되는 유전자를 탐색하 기 위한 tester cDNA로 병 처리 ‘93-3-98’를 사용하고 대조 군으로 사용되는 driver cDNA는 병 처리 ‘스위스트킨’을 사 용하였으며 라이브러리 제작 후 제한효소
RsaI를 처리하였다. 두 개의 튜브에 tester cDNA를 각각 넣고 한 쪽에는 adaptor 1, 다른 한 쪽에는 adaptor 2R을 넣은 후 DNA ligase를 처리 하였다. 각각의 tester pool에 driver cDNA를 첨가하여 hybridization 과정을 거친 다음 차별 발현된 염기서열만 선 택적으로 증폭하기 위하여 2번의 PCR과정을 거쳤다.
비록 SSH가 차등 발현된 전사체를 탐색하기 위한 강력하 고 일반적인 방법으로 사용되지만 SSH 유래 라이브러리는 전형적으로 비 차등적 발현 전사체도 포함되어 background 를 제거하기 위하여 MOS를 실시하였다. SSH의 second nested PCR로부터 얻은 PCR 산물을 phenol/chloroform 처 리를 통하여 추출하였으며 MOS를 수행하기 위하여 에탄올 농축을 실시하였다. PCR 추출물은 제한효소 XmaI 처리를 한 후 denaturation과 5시간의 hybridization 과정을 거친 후 NP2Rs primer를 이용하여 PCR를 수행하였으며 NP2R adaptor 를 양쪽 끝에 연결하였다. MOS PCR 산물을 TOPO TA cloning vector에 삽입하여 형질 전환하였으며 agar, X-gal, IPTG와 ampicillin 100mg/mL 첨가된 LB plate에 도말하여 37°C에서 16시간 배양하였다. 각각의 흰색 colony를 무작위 로 선발하였으며 ampicillin 100mg/mL 첨가된 LB 액체배지 에서 배양한 후 분석에 사용하였다. 각각의 라이브러리 set은 Macrogen Inc. (Seoul, Korea)에서 염기서열을 분석하였다.
cDNA 염기서열 분석
M13 promotor primer (5-CTGGCCGTCGTTTTAC-3)를 사용하여 DNA 염기서열을 분석하였으며 ABI BigDye (R) Terminator v3.1 Cycle Sequencing kit (Applied Biosystems, Lincoln, CA, USA)와 DNA Engine Tetrad 2 Peltier Thermal Cycler (Bio-Rad, Hercules, CA, USA)를 사용하여 분석하 였다. Fluorescent-labeled fragment은 Applied Biosystems 방법에 따라 정제되었으며 ABI 3730xl DNA Analyzer (Applied Biosystems, Lincoln, CA, USA)에서 전기영동을 실시하였으며 염기서열을 분석하였다. database 유사성 분석은 BLASTX 알고리즘(
http://www.ncbi.nlm.nih.gov/)을 이용하 였으며 best matches 염기서열은 similarity scores, E-values 와 annotations을 바탕으로 선택되었다.
SSH 라이브러리 제작에 사용된 동일한 RNA 샘플을 사용 하여 cDNA합성 후 분석하였으며 분석에 사용된 primer는 PRIMER3 (Rozen & Skaletsky, 1998)프로그램을 이용하여 증폭산물의 크기는 100-150bp, annealing온도 60°C로 제작 하였다(Table
1). Endogenous control로써 housekeeping gene인 배 actin (forward: 5-GTGCTGGACTCAGGTGATGG-3, reverse: 5-GTTCTTCTCAACTGACGA GC-3)을 사용하였 으며(
Kim et al. 2009) standard cuve 검정을 통해 안정적으 로 발현되는 것을 확인하였다. Real-time PCR 반응은 총량 20 μl에 SYBR
® Premix Ex Taq™ (Perfect real-time) kit (Takara, Otsu, Shiga, Japan) 10 μl, 0.5 mM의 primer, 20x 희석된 first-strand cDNA, ddH
2O를 혼합하여 최초 95°C에 서 15초간 반응시키고, 95°C에서 5초와 60°C에서 45초간의 cycle를 45회 하여 실시하였으며 마지막 반응으로써 60°C에 서 95°C까지 0.5°C씩 온도를 상승시켜 melting curve분석을 수행하였다. RT PCR은 LightCycler
®480 II (Roche, Rotkreuz, Zug, Switzerland)를 사용하여 3회 반복 분석하였으며 positive calibrate는 무처리 ‘스위트스킨’ 잎 cDNA가 사용되었다. 통 계적 분석 결과는 T-test를 통해 피평가 되었고, 통계적 유의 성은 P<0.05로 정하였다.
Table 1.Quantitative real-time PCR primers used for the relative expression analysis of the ESTs identified by SSH from ‘93-3-98’ pear leaves harvested at 24 and 48hr after inoculation with V. nashicola.
결과 및 고찰
Subtracted cDNA 라이브러리 제작
24H와 48H에서 각각 146과 136개의 clone을 무작위로 선 발하여 염기서열을 분석하였다(Table
2). BLASTX 분석 결 과 database와 유사성을 가지지 못하는 unmatched cDNA의 비율은 24H에서 5.5%, 48H에서 2.9%였으며 두 처리군에서 차등적으로 발현되는 유전자를 비교하였을 때 9개의 유전자 가 24H에서만 발현되었고 14개의 유전자가 48H에서 특이적 으로 발현되었다.
Table 2.Characteristics of SSH libraries from ‘93-3-98’ pear leaves harvested at 24 and 48 hr after inoculation with V. nashicola.
Table 2.
|
Inoculation group |
No. of clones sequenced |
No. of BLASTX matched cDNA |
No. of BLASTX unmatched cDNA |
No. of differentially expressed clones |
|
|
24H |
146 |
138 |
8 |
9 |
|
28H |
136 |
132 |
4 |
14 |
차등 발현 유전자의 염기서열 및 기능 분석
각각의 cDNA 라이브러리의 염기서열을 분석한 결과, 24H 와 48H에서 20개와 26개의 단일 유전자들이 차등 발현되었 으며 방어와 스트레스에 관련된 유전자의 비율은 각각 40% 와 42%였다(Table
1). 사과의 경우 검은별무늬병에 대한 저 항성 품종과 감수성 품종간의 SSH분석을 통해 본 실험과 유 사한 40%의 비율을 보고하였다(
Degenhardt et al. 2005). 식 물의 방어반응인 systemic acquired resistance (SAR)에 관련 된 PR protein family 유전자로써 24H에서는 pathogenesisrelated protein 1a, major allergen Pyr c 1과 major allergen mal d 1, 48H에서는 major allergen mal d 1과 major allergen Mal d 1.03B 유전자가 차등적으로 발현되었으며, 방어반응에 관련된 유전자로는 24H에서 beta-glucosidase, flavonol synthase/ flavanone 3-hydroxylase-like, acyl transferase/acyl hydrolase/ lysophospholipase-like protein, catalase, flavoprotein wrbA, 48H에서 sesquiterpene cyclase, F-box/kelch-repeat protein, peptide transporter, dihydroflavonol-4-reductase isoform 2, cysteine protease inhibitor, glutathione transferase GST 23, 2-oxoglutarate-dependent dioxygenase, cysteine protease, GMFP5 이였다.
‘93-3-98’에
V. nashicola를 접종하여 차등적으로 높게 발현 되는 유전자를 탐색하기 위해 24시간 병 접종 후 샘플을 채취하 였으며 real-time PCR을 이용하여 유전자의 발현을 조사하였다 (Table
3). 유전자의 발현을 분석한 결과 beta-glucosidase, unknown, sesquiterpene cyclase를 제외한 ESTs에서 24시간 병 접종 후 유전자의 발현이 증가하는 것을 보여주었으며, 특히 46개의 DEGs중에서 ripening-induced protein, hypothetical protein POPTRDRAFT_783792, major allergen mal d 1, flavoprotein wrbA, unnamed protein product, hypothetical protein VITISV_037464, hypothetical protein POPTRDRAFT_ 783792, conserved hypothetical protein, 2-oxoglutaratedependent dioxygenase의 9개 유전자가 특이적으로 높게 발 현되었다. 대부분의 PR protein 유전자는 24시간 병 접종 후 발현이 증가하였으며, major allergen Pyr c 1의 경우 배 검 은별무늬병 저항성 품종 ‘황실리’에
V. nashicola를 96시간 접종하여 무처리 군과 유전자의 발현을 비교하였을 때 1.9-fold 발현이 증가 되었지만(
Shin et al. 2012) 본 실험에서는 28.79-fold의 높은 발현차이를 보여주어 조사 품종의 저항성 정도와 접종 후 RNA 추출시기가 이 유전자 발현이 영향하는 것을 확인하였다.
Table 3.Relative expression of selected putative ESTs from ‘93-3-98’ pear leaves harvested at 24 hr after inoculation with V. nashicola. Quantitative real-time PCR was performed to measure the relative expression of each transcript of the ESTs. Actin gene was used as an endogenous control and the data were calibrated relative to the transcript levels in cDNA from uninoculated ‘93-3-98’ pear leaves.
선발된 ESTs의 품종간 발현 비교분석
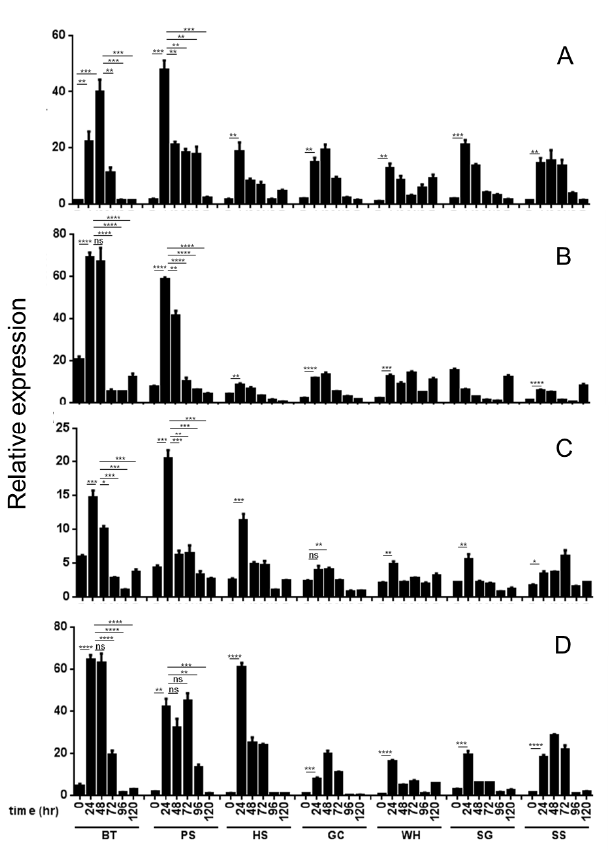
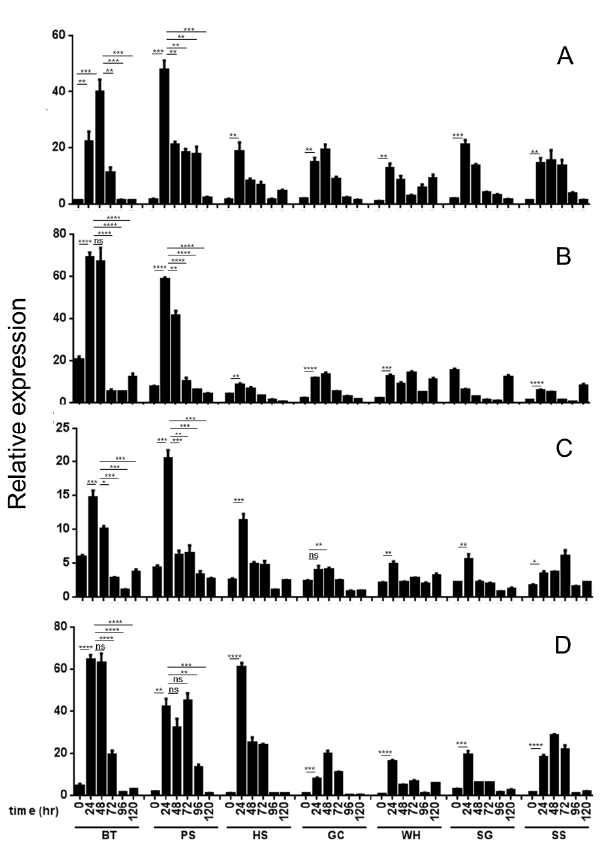
선행된 real-time PCR 결과를 바탕으로 5배 이상 높은 발 현을 가지는 27개의 유전자를 선발하여 고도 저항성 품종 ‘Bartlett’부터 고도 감수성 품종 ‘스위트스킨’까지 7품종을 대상으로 접종시간(24, 48, 72, 96, 120시간)에 따른 유전자 의 발현을 조사한 결과 major allergen Pyr c 1, F-Box/kelchrepeat protein, flavoprotein wrbA, hypothetical protein POPTRDRAFT_783792 유전자에서 저항성 품종 특이적 발 현 차이를 보여주었다. Major allergen Pyr c 1은 모든 품종 에서 24시간 병 접종 후 발현이 높아지는 것을 보여주었는데 특히 고도 저항성 품종인 ‘93-3-98’에서 가장 높은 발현을 보 여주었고(Fig.
1A) 높은 발현을 가지는 고도 저항성 품종 ‘Bartlett’의 경우 접종 24시간 처리에서 발현이 증가하다가 48시간 처리에서 가장 높은 발현을 보였으며 이후 급격히 감 소하였다. 따라서 major allergen Pyr c 1은 고도 저항성 품 종에서 높게 발현되며 병 접종 초기에 발현이 증가하는 경향 을 보였다.
Fig. 1.Relative expression of Major allergen Pyr c 1 (A), F-box/kelch-repeat protein (B), Flavoprotein wrbA (C) and hypothetical protein (D) genes in various pear cultivars having different scab-susceptibilities at 0, 24, 48, 72, 96 and 120 h after inoculation with V. nashicola. BT, ‘Bartlett’; PS, ‘93-3-98’; HS, ‘Hwangsilri’; GC, ‘Gamcheonbae’; WH, ‘Wonhwang’; NI, ‘Niitaka’; SS, ‘Sweat Skin’. Data are expressed as mean +/- standard error (A-D, n=3). *P < 0.05; **P < 0.01; ***P < 0.001, ****p < 0.0001. Quantitative real-time PCR was performed to measure the relative expression of each transcript of the ESTs. Actin gene was used as an endogenous control and the data were calibrated relative to the transcript levels in cDNA from uninoculated ‘Sweat Skin’ pear leaves.
모든 식물들은 병원체로부터 공격을 받았을 때 다양한 방 어 유전자들이 발현된다. 그 중 대표적인 단백질 중의 하나가 PR protein으로(
Van Loon, 1985) SAR 메커니즘에 중요하 게 관련되어 있다. PR protein은 구조적 특징과 생물학적 활성 에 기초하여 17가지의 family로 구분되며 beta-1,3-glucanase, chitinase, thaumatin-like protein, peroxidase, ribosomeinactivating protein, defense, thionins, nonspecific lipid transfer proteins, oxalate oxidase, oxalate-oxidase-like protein, protease inhibitor 등이 포함되어 있다(
Van Loon & Van Strien 1999). 이들 중 chitinase와 β-1,3-glucanases는 중요 한 가수분해 효소로써 특이적 가수분해 활성화와 세포벽의 파괴 등을 통해 균류의 생장을 저해할 수 있는 것으로 알려져 있다(
Mauch et al. 1988). Major allergen Pyr c 1은 PR10 Group에 속하여(
Lebel et al. 2010) 상처(
Graham et al. 2003), 병원체 감염(
Park et al. 2004), 발아(
Leubner-Metzger 2002), 물리적 스트레스(
Dat et al. 2000) 등에 의해 발현이 유도되어 진다. PR10은 ribonuclease 활성을 가지며(
Filipenko et al. 2013) 환경적인 스트레스나 병원체에 의해 공격을 받았을 때 거의 모든 기관 및 조직에서 유전자 발현이 급격히 증가함에 따라 방어반응에 중요하게 관여하는 것으로 인식되고 있다 (
Liu & Ekramoddoullah 2006).
Major allergen Pyr c 1과 마찬가지로 F-Box/kelch-repeat protein 유전자의 발현을 분석한 결과 다른 품종에 비해 ‘Bartlett’에서 가장 높게 발현되었으며 다음으로 ‘93-3-98’에 서 높았다(Fig.
1B). 병 접종 시간에 따른 발현양상은 24시간 처리구에서 높게 발현 되었으며 48시간 처리구에서 다소 감 소하다가 72시간 이후 급격히 감소하였다.
F-box protein은 최소 하나 이상의 F-box domain을 포함 하는 단백질로 Skp, Cullin, F-box containing complex (SCF complex)의 한 구성 분자로 최초에 동정되었으며(
Schulman et al. 2000) F-box domain은 특징적으로 protein-protein interaction에 관여하여 다양한 기질 단백질과 결합을 통해 특 정 단백질의 ubiquitination을 유도함으로써 proteosome에서 단백질 분해에 작용한다. 이를 통해 단백질의 안정성 조절과 cell cycle 조절 및 다양한 세포내 신호전달에 관여하고 있다 (Craig & Tyers 1999). 뿐만 아니라 식물호르몬 신호전달에 관여함으로써 식물 세포의 분화 과정이나 스트레스 반응에서 도 중요하게 작용하고 있음이 보고되고 있다(
Yan et al. 2011). 최근 식물에 있어서 protease와 더불어 ubiquitin을 통 한 단백질 파괴가 스트레스 환경뿐만 아니라 병 저항성에 관 여하고 있음이 많이 보고되고 있다. 쌀에서 많이 발현되는 F-box protein 중의 하나인
OsDRF1의 과 발현에 의해 병 저 항성이 증가됨이 보고되었고(
Cao et al. 2008), 병원체 인식 에 중요하게 작용하는 수용체 중의 하나인 NLR의 활성도 F-box protein에 의한 ubiquitination에 의해 조절 받고 있음 이 보고되었다(
Cheng et al. 2011). 자세한 기작에 대한 연구 는 더 필요하겠지만, F-box protein에 의한 방어 메커니즘은 독성에 관여하는 단백질을 ubiquitin화 시켜 분해함으로써 독 성에 대한 저항성을 나타낼 것으로 기대되고 있다(
Hwang et al. 2009).
Flavoprotein wrbA은 고도 저항성 품종 ‘Bartlett’, ‘93-3-98’ 에서 24시간 병 접 종 이후 높은 발현을 보였고 저항성 품종 ‘황실리’ 또한 감수성 품종과 비교하여 24시간 병 접종 처리 구에서 높은 발현을 보였다(Fig.
1C). Flavoprotein wrbA은 다양한 퀴논에 의해 NAD(P)H를 NAD(P)
+로 산화시킴으로 써 활성산소를 제거하는 효소로 최근 염 저항성 잔디품종에 서 특이적으로 발현됨이 보고되었다(
Liu et al. 2012).
Hypothetical protein 또한 다른 품종에 비해 고도 저항성 품종 ‘Bartlett’, ‘93-3-98’에서 24시간 병 접종 이후 높은 발현 을 보였으며, 저항성 품종 ‘황실리’에서도 고도 저항성 품종과 비슷한 높은 발현을 보였다(Fig.
1D). 이 결과는 hypothetical protein의 정확한 기능을 확인할 수 없으나 저항성 품종에서 특이적으로 발현되는 것으로 보아 저항성에 관련되어 있을 것이라 보여진다.
Major allergen Pyr c 1, F-Box/kelch-repeat protein, flavoprotein wrbA, hypothetical protein 유전자가 배 검은별무늬 병에 관련하여 어떠한 역할을 하는지 정확히 알 수 없다. 하 지만 앞으로 배 검은별무늬병 감수성 품종의 형질전환체계를 확립하여 이러한 유전자를 도입한 후 표현형과 유전자의 발 현을 분석한다며 좀더 구체적인 기능을 밝힐 수 있을 것이라 판단된다.
초 록
본 연구는 V. nashicola에 의해 발생하는 배 검은별무늬병 에 저항성을 가지는 유전자를 선발하고자 고도 저항성 ‘93-3-98’과 고도 감수성 ‘스위트스킨’에 병 접종후 24시간, 48시간후 엽을 채취하여 suppression subtractive hybridization 을 수행하였다. ESTs 분석결과 ‘93-3-98’에서만 발현된 유전 자는 24H(tester, 24시간 ’93-3-98; driver, 24시간 ‘스위트스 킨’)과 48H(tester, 48시간 ’93-3-98; driver, 48시간 ‘스위트 스킨’)에서 각각 9개, 14개였으며 방어 또는 스트레스에 관련 된 유전자의 비율은 각각 40%와 42%였다. 식물의 방어반응에 관련된 PR protein family 유전자로써는 24H에서 pathogenesisrelated protein 1a, major allergen Pyr c 1과 allergen mal d 1, 48H에서는 major allergen Mal d 1.03B 유전자가 특이 적으로 발현되었고 다수의 방어반응에 관련된 유전자들도 확 인되었다. Major allergen Pyr c 1, F-Box/kelch-repeat protein, flavoprotein wrbA, hypothetical protein POPTRDRAFT_ 783792은 고도저항성 ‘Bartlett’과 ‘93-3-98’에서만 높게 발 현 되었으며 중도저항성 ‘감천배’와 감수성 ‘원황’, 고도 감수 성 ‘신고’와 ‘스위트스킨’에서는 낮게 발현되어 검은별무늬병 저항성 연관 유전자로 확인되었다.
사 사
본 연구는 농촌진흥청 연구사업(과제번호: PJ0065952013) 의 지원에 의해 이루어진 것임.
References
- 1. Bouvier L, Bourcy M, Boulay M, Tellier M, Guérif P, Denancé C, Durel CE, Lespinasse Y. A new pear scab resistance gene Rvp1 from the European pear cultivar ‘Navara’maps in a genomic region syntenic to an apple scab resistance gene cluster on linkage group 2. Tree Genet. Genomes 2012. 8: 53-60.
- 2. Cao Y, Yang Y, Zhang H, Li D, Zheng Z, Song F. Overexpression of a rice defense-related F-box protein gene OsDRF1 in tobacco improves disease resistance through potentiation of defense gene expression. Physiol. Planntarum 2008. 134: 440-452.
- 3. Cheng YT, Li Y, Huang S, Huang Y, Dong X, Zhang Y, Li X. Stability of plant immune-receptor resistance proteins is controlled by SKP1-CULLIN1-F-box (SCF)-mediated protein degradation. P. Natil. Acad. Sci 2011. 108: 14694-14699.
- 4. Cho KH, Shin IS, Kim KT, Suh EJ, Hong SS, Lee HJ. Development of AFLP and CAPS markers linked to the scab resistance gene, Rvn2, in an inter-specific hybrid pear (Pyrus spp). Hort. Sci. Biotechnol 2009. 84: 619-624.
- 5. Dat J, Vandenabeele S, Vranova E, Van Montagu M, Inzé D, Van Breusegem F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life. Sci 2000. 57: 779-795.
- 6. Degenhardt J, Al-Masri AN, Kürkcüoglu S, Szankowski I, Gau AE. Characterization by suppression subtractive hybridization of transcripts that are differentially expressed in leaves of apple scab-resistant and susceptible cultivars of Malus domestica. Mol. Genet. Genomics 2005. 273: 326-335.
- 7. Faize M, Faize L, Ishizaka M, Ishii H. Expression of potential defense responses of Asian and European pears to infection with Venturia nashicola. Physiol Mol. Plant Pathol 2004. 64: 319-330.
- 8. Filipenko E, Kochetov A, Kanayama Y, Malinovsky V, Shumny V. PR-proteins with ribonuclease activity and plant resistance against pathogenic fungi. Russ. J. Genet Applied Research 2013. 3: 474-480.
- 9. Graham M, Weidner J, Wheeler K, Pelow M, Graham T. Induced expression of pathogenesis-related protein genes in soybean by wounding and the Phytophthora sojae cell wall glucan elicitor. Physiol Mol. Plant Pathol 2003. 63: 141-149.
- 10. Hammond-Kosack KE, Jones J. Resistance genedependent plant defense responses. Plant Cell 1996. 8: 1773.
- 11. Hwang GW, Wada N, Kuge S, Naganuma A. Overexpression of the novel F-box protein Ymr258c confers resistance to methylmercury in Saccharomyces cerevisiae. J. Toxicol. Sci 2009. 34.
- 12. Iandolino A, da Silva FG, Lim H, Choi H, Williams L, Cook D. High-quality RNA, cDNA, and derived EST libraries from grapevine (Vitis vinifera L). Plant Mol. Biol. Rep 2004. 22: 269-278.
- 13. Ishii H, Udagawa H, Nishimoto S, Tsuda T, Nakashima H. Scab resistance in pear species and cultivars. Acta. Phytopathol. Hun 1992. 27: 293-298.
- 14. Kim SH, Koh GC, Hwang B, Lee HJ, Hong SW. Identification of differential gene expression in juvenile vs mature leaves of pear (Pyrus pyrifolia) by using annealing control primer. J. Korea. Soc. Appl. Biol. Chem 2009. 52: 121-127.
- 15. Lebel S, Schellenbaum P, Walter B, Maillot P. Characterisation of the Vitis vinifera PR10 multigene family. BMC Plant Biol 2010. 10: 184.
- 16. Leubner-Metzger G. Seed after-ripening and overexpression of class I ?-1, 3-glucanase confer maternal effects on tobacco testa rupture and dormancy release. Planta 2002. 215: 959-968.
- 17. Liu JJ, Ekramoddoullah AK. The family 10 of plant pathogenesis-related proteins their structure, regulation, and function in response to biotic and abiotic stresses. Physiol. Mol. Plant Pathol 2006. 68: 3-13.
- 18. Liu Y, Du H, He X, Huang B, Wang Z. Identification of differentially expressed salt-responsive proteins in roots of two perennial grass species contrasting in salinity tolerance. J Plant Physiol 2012. 169: 117-126.
- 19. Mauch F, Hadwiger LA, Boller T. Antifungal hydrolases in pea tissue I Purification and characterization of two chitinases and two β-1, 3-glucanases differentially regulated during development and in response to fungal infection. Plant Physiol 1988. 87: 325-333.
- 20. Moody D. Genomics techniques An overview of methods for the study of gene expression. J. Anim. Sci 2001. 79(Suppl): E1280-E1135.
- 21. Pan D, He N, Yang Z, Liu H, Xu X. Differential gene expression profile in hepatopancreas of WSSV-resistant shrimp (Penaeus japonicus) by suppression subtractive hybridization. Dev. Comp. Immunol 2005. 29: 103-112.
- 22. Paparu P, Dubois T, Coyne D, Viljoen A. Defenserelated gene expression in susceptible and tolerant bananas (Musa spp) following inoculation with non-pathogenic Fusarium oxysporum endophytes and challenge with Radopholus similis. Physiol. Mol. Plant Pathol 2007. 71: 149-157.
- 23. Park CJ, Kim KJ, Shin R, Park JM, Shin YC, Paek KH. Pathogenesis-related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral pathway. Plant. J 2004. 37: 186-198.
- 24. Park P, Ishii H, Adachi Y, Kanematsu S, Ieki H, Umemoto S. Infection behavior of Venturia nashicola, the cause of scab on Asian pears. Phytopathology 2000. 90: 1209-1216.
- 25. Schulman BA, Carrano AC, Jeffrey PD, Bowen Z, Kinnucan ER, Finnin MS, Elledge SJ, Harper JW, Pagano M, Pavletich NP. Insights into SCF ubiquitin ligases from the structure of the Skp1-Skp2 complex. Nature 2000. 408: 381-386.
- 26. Shin IS, Bae KM, Nam GY, Kang BC, Chun J, Cho KH, Kim SH, Choi HS, Kim HR, Hwang HS. Identification of genes induced by Venturia nashicola in indigenous Korean pear 'Hwangsilri'. Hort. Environ. Biotechnol 2012. 53: 513-520.
- 27. Sticher L, Mauch-Mani B, M´etraux JP. Systemic Acquired Resistance. Annu. Rev. Phytopathol 1997. 35: 235-70.
- 28. Terakami S, Shoda M, Adachi Y, Gonai T, Kasumi M, Sawamura Y, Iketani H, Kotobuki K, Patocchi A, Gessler C. Genetic mapping of the pear scab resistance gene Vnk of Japanese pear cultivar Kinchaku. Theor. Appl. Genet 2006. 113: 743-752.
- 29. Van Loon L. Pathogenesis-related proteins. Plant. Mol. Biol 1985. 4: 111-116.
- 30. Van Loon L, Van Strien E. The families of pathogenesis- related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol 1999. 55: 85-97.
- 31. Xiong L, Lee MW, Qi M, Yang Y. Identification of defense-related rice genes by suppression subtractive hybridization and differential screening. Mol. Plant Microbe. Interact 2001. 4: 685-692.
- 32. Yan YS, Chen XY, Yang K, Sun ZX, Fu YP, Zhang YM, Fang RX. Overexpression of an F-box protein gene reduces abiotic stress tolerance and promotes root growth in rice. Mol. Plant 2011. 4: 190-197.
- 33. Zheng QL, Ishii H. Molecular cloning and expression analysis of genes related to phosphatidic acid synthesis inJapanese pear leaves inoculated with Venturia nashicola. J. Gem. Plant Pathol 2009. 75: 413-421.