적요
The β-carotene biofortified transgenic soybean was developed recently through Agrobacterium-mediated transformation using the recombinant PAC (Phytoene synthase-2A-Carotene desaturase) gene in Korean soybean (Glycine max L. cv. Kwangan). GM crops prior to use as food or release into the environment required risk assessments to environment and human health in Korea. Generally, transgenic plants containing a copy of T-DNA were used for stable expression of desirable trait gene in risk assessments. Also, information about integration site of T-DNA can be used to test the hypothesis that the inserted DNA does not trigger production of unintended transgenic proteins, or disrupt plant genes, which may cause the transgenic crop to be harmful. As these reasons, we selected four transgenic soybean lines expressing carotenoid biosynthesis genes with a copy of T-DNA by using Southern blot analysis, and analyzed the integration sites of their T-DNA by using flanking sequence analysis. The results showed that, T-DNA of three transgenic soybean lines (7-1-1-1, 9-1-2, 10-10-1) was inserted within intergenic region of the soybean chromosome, while T-DNA of a transgenic soybean line (10-19-1) located exon region of chromosome 13. This data of integration site and flanking sequences is useful for the biosafety assessment and for the identification of the β-carotene biofortified transgenic soybean.
서 언
유전자변형(Genetically modified, GM) 기술을 이용한 제 초제내성 GM콩(RRS, Roundup Ready Soybean)이 1996년 개발되어 상업적 성공을 거둔 이래 GM작물은 꾸준한 성장세 를 나타내고 있다. GM작물은 2014년 기준 전세계 약 1억 8,150만 헥타르 면적에서 재배되고 있으며(
James 2013), 국 내의 경우 재배 승인을 얻어 재배되는 GM작물은 없지만 7작 물 127종이 수입승인 되었고(2015년 3월 현재기준), 2014년 에는 약 31억 달러(1,100만 톤)의 GM작물이 식용 및 사료용 으로 수입되었다(
KBCH 2015). 이중 콩(대두)은 국내에 수입 되고 있는 GM작물 중 옥수수와 함께 가장 많이 수입되고 있 는 작물로 식용유 생산과 가축의 단백질 공급을 위한 사료 제 조에 활용되고 있다. 현재까지 승인 받은 GM콩 품종은 제초 제내성 형질을 갖도록 개발된 품종과 후대 교배종들이 대부 분을 차지하지만, 최근에는 올레인산(Oleic acid) 증진된 DP 305423및 스테아리돈산(Stearidonic acid) 생성 유전자가 도 입된 MON87769의 경우와 같이 지방산 성분을 강화시킨 품 종들이 증가하고 있다.
카로티노이드는 자연계에 널리 분포하는 중요 색소물질로 서 식물은 물론 세균, 미세조류, 곰팡이에서도 합성되며, 식물 에서는 엽록체(Chloroplast)와 유색체(Chromoplast)에서 생 합성 되며(
Ha et al. 2007), 인체에서는 비타민 A의 전구물질 로 이용되며 항산화 효과가 뛰어나 노화방지, 면역증진 등에 효과가 있는 것으로 보고된 바 있다(
Stahl & Sies 2003). 작 물에서 대사공학을 이용하여 카로티노이드 함량을 높이려는 연구는 벼에서 먼저 시작되었다. 저개발 국가의 비타민A 결 핍문제를 해결하기 위한 일환으로
Japonica 쌀 품종에 수선 화 유래의 phytoene synthase (
Psy) 유전자와 세균(
Erwinia uredovora) 유래의 carotene desaturase (
CrtI) 유전자를 도입 해 베타카로틴 함량을 증진시킨 유전자변형 벼(Golden Rice) 가 최초로 개발되었고(
Ye et al. 2000), 국내에서도 낙동벼 (
Oryza sativa L. cv. Nakdong)에 녹광 고추(
Capsicum annuum cv. Nockwang)에서 분리한
Psy 유전자와
CrtI 유전자를 2A 다중발현 시스템으로 발현 유도하여 베타카로틴 함량을 높인 비타민A 강화 벼가 개발된 바 있다(
Ha et al. 2010).
최근 국내 콩 품종인 광안(
Glycine max L. cv. Kwangan) 을 이용하여 비타민A 강화 벼 개발에 이용된
Psy와
CrtI 유 전자를 종자 특이적 β-conglycinin 프로모터로 발현 유도해 카로티노이드 함량이 비형질전환 모본 대비 약 62배 증진된 형질전환 콩이 개발되었다(
Kim et al. 2012). 유전자변형작물 의 활용을 위해서는 도입유전자의 기능 검정 뿐 아니라 개발 단계부터 식품 및 환경에 대한 위해성평가 과정을 거쳐 안전 성을 검증하여야 하며 GM작물의 분자생물학적 평가정보는 위해성 평가의 필수 요소이다. 본 연구에서는 베타카로틴 강 화 콩의 안전성평가를 위한 재료확보 및 기초자료로 활용하 고자 분자생물학적 방법으로 단일 도입유전자(T-DNA)가 삽 입된 후대계통을 선발하고 도입유전자의 삽입위치를 분석하 였다.
재료 및 방법
식물재료
본 실험의 재료는 2A 다중발현 시스템을 사용하여 종자 특 이적 β-conglycinin 프로모터에 의해 카로티노이드 합성 관 련
Psy와
CrtI 유전자가 공동 과발현 되도록 개발된 binary vector pB2GW7.0-β-conglycinin (Fig.
1)를 애그로박테리움 을 이용해 국내품종인 광안콩에 형질전환하여 개발된 베타카 로틴 강화 콩이며(
Kim et al. 2012), 대조품종으로는 비형질 전환 모본인 광안 품종이 사용되었다.
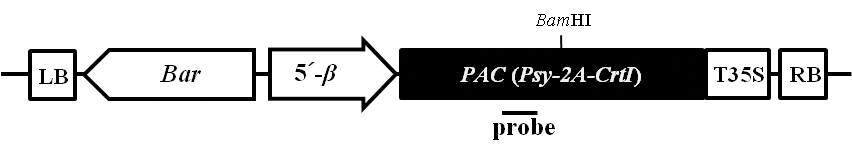
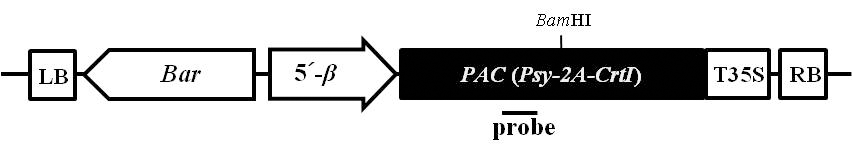
Fig. 1.Schematic diagram of the T-DNA region of binary vector used in the soybean transformation. LB and RB, left and right borders for Agrobacterium-mediated transformation, respectively; Bar, the DL-phosphinothricin resistance gene driven by the nopaline synthase (nos) promoter and nos terminator; 5´-β, a soybean seed specific β-conglycinin promoter; PAC, a recombinant Psy-2A-Tp-CrtI gene; T35S, a cauliflower mosaic virus 35S terminator. The probe used for Southern blot analysis was marked by black line.
단일 도입유전자가 도입된 후대계통 선발을 위해서 T2세대 이후의 형질전환 콩 중 비교적 농업적 형질이 양호한 계통을 선발하여 분석에 활용하였으며, 실험의 대조체로는 비형질전 환 모본인 광안 품종이 사용되었다.
형질전환 작물의 도입유전자 수 결정
형질전환 베타카로틴 강화 콩에 삽입된 도입유전자의 수 결정은 서던 분석을 사용하였다. 분석을 위한 genomic DNA 추출은 CTAB 방법을(
Doyle & Doyle 1987) 이용하였으며 추출된 genomic DNA는 NanoDrop spectrophotometer ND- 1000 (NanoDrop technologies Inc., Wilmington, DE, USA) 를 이용하여 정량 하였다. 분리된 식물 genomic DNA는 제 한효소
BamHI으로 완전절단하여 0.8% 아가로스 겔에 전기 영동하였다. 서던 분석을 위한 probe는
Psy 유전자 특이적 primer (Table
1)을 이용하여 PCR DIG probe synthesis kit (Roche, Florence, SC, USA)로 합성하였고, hybridization 및 detection 방법은
Qin et al. (2013)이 기술한 방법에 따라 수행하였다.
Table 1.Sequences of primers used in this study.
Table 1.
|
Name |
Primer sequence (5′–3′) |
Specificity |
|
|
Psy-for |
ATTACTCCGGCGGCCTTAGA |
Psy
|
|
Psy-rev |
TCGGATAGACCTGCCTGTGC |
|
|
Pac-for-1 |
ATGTCTGTTGCCTTGTTATGG |
PAC (Psy-2A-CrtI)
|
|
Pac-rev-4 |
CAAATCAGATCCTCCAGCAT |
|
|
Lectin F |
CTTGGTCGCGCCCTCTACTC |
Lec
|
|
Lectin R |
AAGGCAAGCCCATCTGCAAG |
|
|
Ada1 |
GCGTAATACGACTCACTATAGCAATTAACC |
Adaptor |
|
Ada2 |
GACTCACTATAGCAATTAAC |
|
|
LB1 |
CGGCTATTGGTAATAGGACACTGG |
LB region |
|
LB2 |
CTGCTGAAGTCCCTGGAGGC |
|
|
RB1 |
CTGATACCAGACGTTGCCCGCATAA |
RB region |
|
RB2 |
CCTGGCTTACGGAGTCAGCT |
|
|
7-1-1-1-F |
CAGATCATGGCCAGAACACGAGG |
Flanking region of 7-1-1-1 |
|
7-1-1-1-R |
AGTCACGCACTGATGCACACTAG |
|
|
9-1-2-F |
TGGCCAAGAGGTTTAGCGTCCT |
Flanking region of 9-1-2 |
|
9-1-2-R |
GGGCGAGAGTTTATATTGCCATT |
|
|
10-19-1-F |
AGTCTGGGAAGCGAAAGCTTGG |
Flanking region of 10-19-1 |
|
10-19-1-R |
GATGTGGTTTCATCCATTGCTAGCTG |
|
|
10-10-1-F |
ACAAACACGGATTGGTCCAAG |
Flanking region of 10-10-1 |
|
10-10-1-R |
TAGCGTGCCACATGGAAGAGCGAAA |
|
도입유전자 확인을 위한 PCR, RT-PCR 및 immunostrip 검정
형질전환 콩에 삽입된 도입유전자의 발현 분석은 RT-PCR 방법으로 수행되었다. 분석에 사용된 total RNA의 추출은 Qiagen 사(Qiagen, Germantown, MD, USA)의 RNeasy Plant mini kit (USA)를 이용하였다. 분리된 total RNA는 DNase 처리후 RevertAid
TM First strand cDNA synthesis kit (Thermo Scientific, Waltham, MA, USA)로 cDNA로 변환하여 PCR 증폭의 template로 사용하였다. PCR 증폭은 250 ng cDNA, 10× Ex Taq Buffer 5 μl (Takara Inc., Otsu, Shiga, Japan), dNTP mixture (2.5 mM each) 4 μl, Ex Taq polymerase (2.5 unit) 0.5 μl, sense primer (20 pmoles) 1μl, anti-sense primer (20 pmoles) 1 μl를 각각 첨가한 전체 50 μl의 반응 물을 95°C에서 5 분간 전 처리 후, 98°C 10초, 58°C 30초, 72°C 3분을 1주기로 30회 연속 반응하고 72°C에서 10분간 후 처리하여 수행하였다. RT-PCR 분석의 내부 표준 유전자 로는 콩의 내재 lectin 유전자가 이용되었고 분석에 사용된 primer는 Table
1에 나타내었다. 도입유전자 확인을 위한 PCR 분석을 위한 total DNA 분리는 서던 분석 방법에 기술 된 CTAB 방법을 사용하였으며 PCR 조건은 RT-PCR에 사 용된 DNA 증폭 조건과 동일하게 수행되었다.
도입유전자의 bar 단백질 발현 확인을 위한 immunostrip 검정(lateral flow strip test)은 시료를 마쇄하고 추출액을 넣 어 단백질을 추출한 후, 제조사가 제공한 방법에 따라 Trait LL Test Strip (Strategic Diagnostics Inc., Newark, DE, USA)을 이용하여 수행하였다.
도입유전자의 삽입 위치 분석
형질전환 콩에서 도입유전자의 삽입위치 확인은
Bugert et al. (2001)이 보고한 PCR-walking 방법을 개량하여 사용하였 다. 실험에 사용된 primer는 Table
1에 나타내었고 구체적 실 험방법은 다음과 같다.
500 ng의 total DNA를 2 U의 HaeIII 제한효소로 완전절 단 시킨 뒤 5 U의 T4 DNA ligase (Takara Inc., Otsu, Shiga, Japan)과 50 pmol의 adaptor DNA를 첨가하여 37°C에서 1 시간 반응시켜 두 단계의 PCR을 수행하였다. 일차 PCR은 전 체 20 μl pre-mixture 반응물 안에 0.5 pmol의 Ada1과 LB1 또는 RB1 primer를 첨가하여 PTC-200 thermal cycler (MJ Research, Waltham, MA, USA)를 이용하여 반응시켰다. 일 차 PCR 반응조건은 95°C에서 5분간 전 처리한 후, 94°C에서 30초, 67°C에서 1분, 72°C에서 1분간을 기본주기로 하여 총 20회 반복 반응하고 72°C에서 10분간 최종 증폭시켰다. 이차 PCR은 일차 PCR 생성물 5 μl을 주형으로 하여 Ada2와 LB2 또는 RB2 primer를 사용하여 94°C에서 5분간 전 처리 한 후 94°C에서 30초, 60°C에서 30초, 72°C에서 1분 30초간 을 반응주기로 전체 40회 반복 반응시킨 후 72°C에서 10분간 최종 증폭하여 진행하였다. 증폭된 PCR 반응산물은 1% agarose gel에 전기영동 하여 확인하고, 증폭된 DNA 밴드는 HiYieldTM Gel/PCR DNA Extraction Kit (RBC Bioscience, Taipei, Taiwan)를 이용하여 정제하였으며 LB2 또는 RB2 primer를 이용하여 ABI3730XL DNA 분석기(Applied Biosystems, FosterCity, CA, USA)로 염기서열 분석하였다. 분 석된 염기서열 정보는 Ensembl 시스템의 Gramene BLAST Search (http://ensembl.gramene.org/Tools/Blast?db=core)와 NCBI의 Blast 프로그램(http://blast.ncbi.nlm.nih.gov/)을 이 용하여 분석되었다.
결과 및 고찰
도입유전자의 복제 수 및 발현 분석
애그로박테리움 형질전환 방법을 사용하여 복수(multiple) 의 T-DNA가 삽입된 6 계통의 형질전환 베타카로틴 강화 콩 이 개발되었다(
Kim et al. 2012). T-DNA의 copy 수와 발현 량의 관계는 일반적으로 형질전환체에 도입된 T-DNA의 copy 수가 많을수록 그 발현량이 높다고 알려져 있으나 정비 례하지 않으며, 단일 copy의 T-DNA 도입이 도입유전자의 안정적 유지와 발현을 위해 유리하다고 보고된 바 있다(
De Buck et al. 2004,
Schubert et al. 2004).
Kim et al. (2012)의 보고에서 베타카로틴 강화 콩 계통 가 운데 베타카로틴 함량이 비교적 높았던 T
1 세대 3 계통(7-3, 13-5, 16-2)의 도입유전자 copy 수는 7번 계통에서 3-4 copy, 13번 계통은 7-8 copy, 및 16번 계통은 2-3 copy로 분석되었 다. 자가수분을 통한 도입유전자 분리(gene segregation)로 단일 copy T-DNA가 도입된 형질전환체를 선발하기 위해 T
2 세대 베타카로틴 강화 콩을 국립농업과학원 GMO온실에 증 식하여 7-3 계통은 T
6 세대까지 13-5, 16-2 계통은 T
5세대까 지 세대 전개하였다. 이후 도입유전자 copy 수 확인을 위해 증식된 T
5, T
6 후대계통을 무작위 선발하여 genomic DNA를 추출하고 형질전환 벡터에 단일 제한효소 자리가 존재하는
BamHI으로 절단한 후
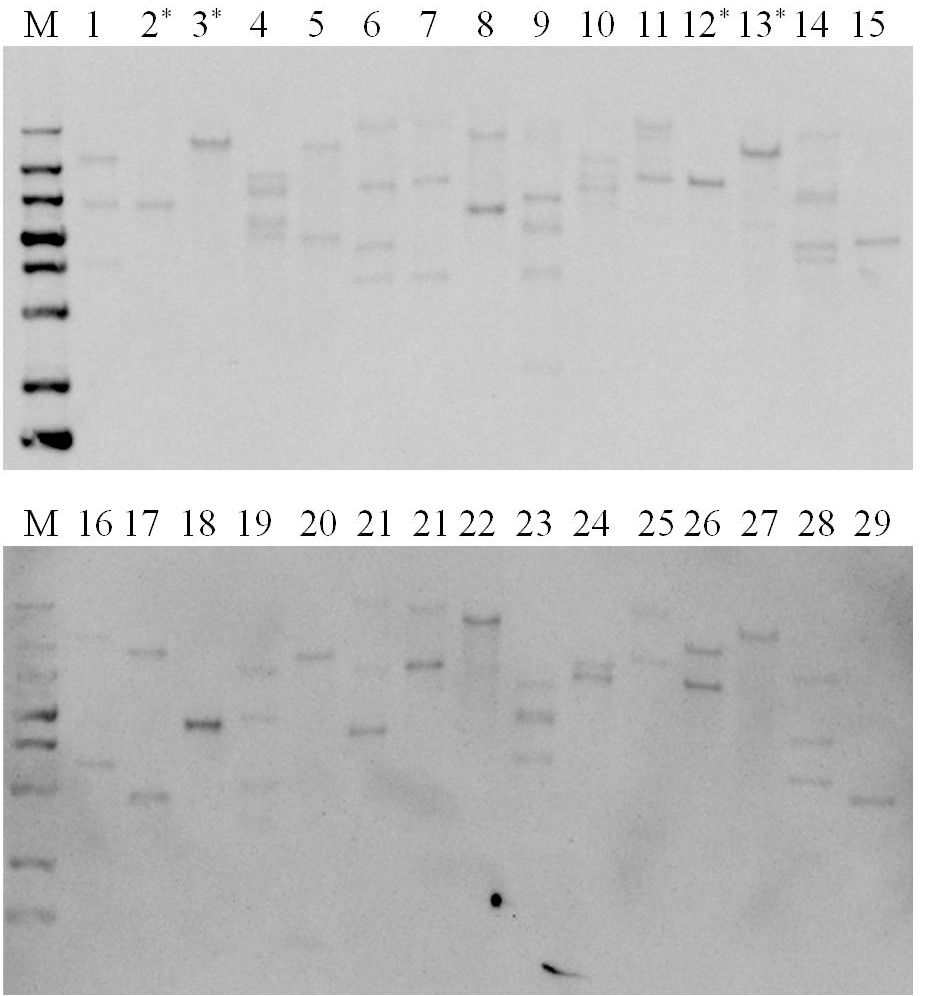
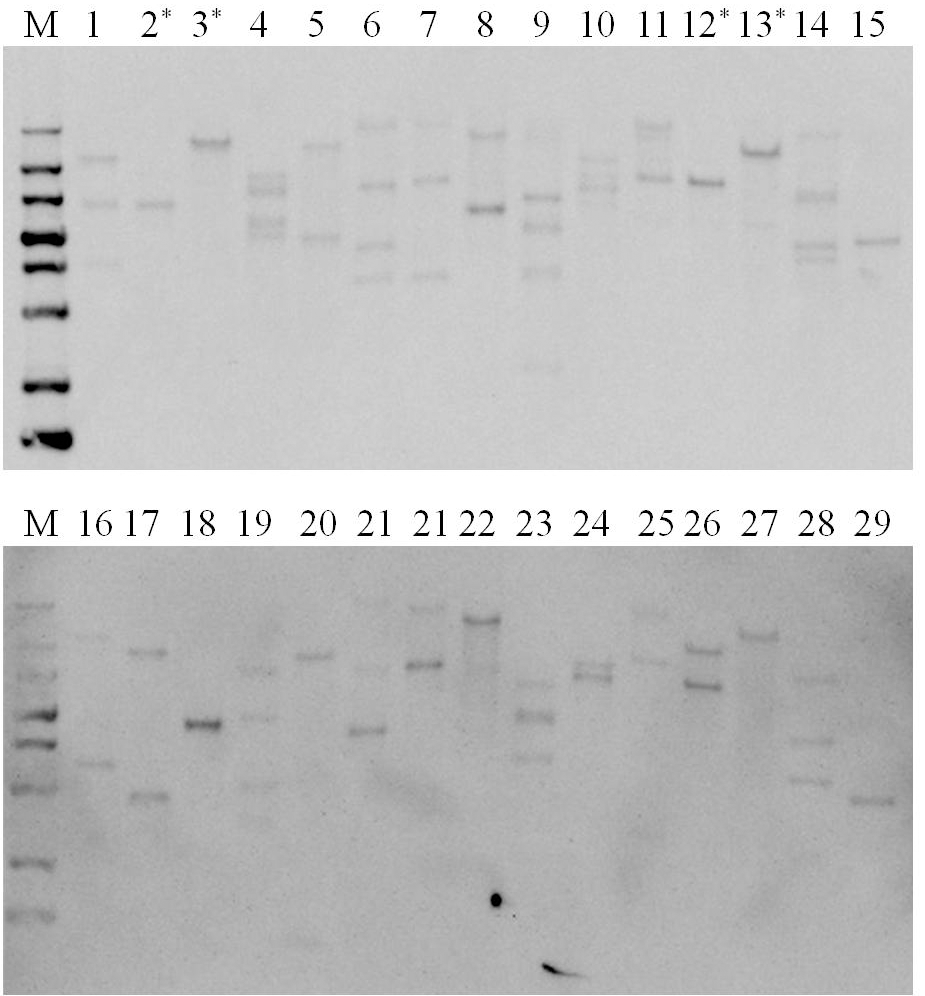
Psy 유전자의 단편을 probe로 사용하 여 서던 분석을 수행하였다. Fig.
2에 나타낸 바와 같이 도입 유전자가 1 copy 에서 5 copy까지 베타카로틴 콩 후대 계통 에서 분리된 것을 확인할 수 있었다. 서던 분석 결과에 기초 해 단일도입유전자 삽입된 것으로 확인된 계통 중 4개의 후대 계통을(Fig.
2의 2, 3, 12, 13 line) 이후의 분석을 위해 선발 하고, 7-1-1-1(T
2 세대 7-3번 형질전환체 유래)과 9-1-2, 10-19-1, 10-10-1(T
2 세대 13-5번 형질전환체 유래)로 각각 명 명하였다.
Fig. 2.Southern blot analysis for copy number confirmation of transgenic soybean lines. M, DIG-labeled molecular marker; lanes 1-29, transgenic soybean lines; Asterisked lines represent selected transgenic soybean lines for further assay.
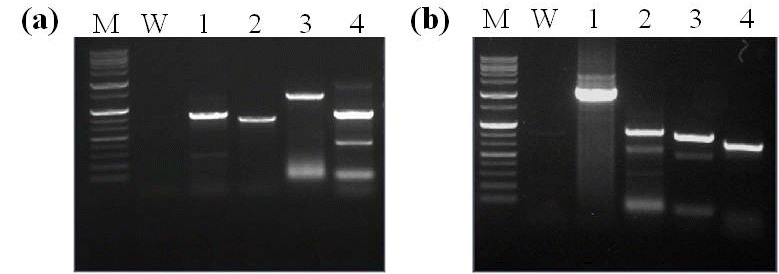
선발된 4개 후대계통들의 도입유전자와 발현 확인을 검정 하기 위해 T-DNA의
PAC (
Psy-2A-CrtI) 유전자 부위를 이용 하여 PCR 및 RT-PCR 분석을 실시하였다. 분석결과 선발된 4계통 모두에서
PAC 유전자가 확인되었고 강하게 발현되는 것을 확인할 수 있었다(Fig.
3a 및
3b). 또한 베타카로틴 강화 콩에
PAC 유전자와 함께 도입된
bar 유전자의 발현 확인을 위한 lateral flow immunostrip 검정에서도 9-1-2 계통에서는 bar 단백질의 밴드 패턴이 다소 약했지만 후대계통 모두에서 bar 단백질의 발현을 확인할 수 있었다(Fig.
3c).
Fig. 3.Identification of the T-DNA in transgenic soybean plants. (a) PCR analysis of transgenic soybean plant lines for identification of T-DNA. (b) RT-PCR analysis of PAC from leaves of transgenic soybean plant lines. The soybean lectin (Lec) gene was used for normalization. (c) immunostrip tests to detect bar protein expressed in transgenic soybean lines. M, molecular marker; W, nontransformed wild-type Kwangan; P, positive control; Lanes 1-4 indicate transgenic soybean line 7-1-1-1, 9-1-2, 10-19-1, and 10-10-1, respectively.

선발된 후대 계통들을 온실에서 생육시킨 결과 10-10-1 계 통은 모본인 광안에 비해 다소 초장이 작았지만 다른 7-1-1-1 및 9-1-2, 10-19-1 계통은 모본과 유사하였다(Fig.
4a), 또한 형질전환 후대계통들의 종자는 모본에 비해 전체적으로 크기 는 다소 작았지만 종자단면은 높은 카로티노이드 함량을 의 미하는 주황색이 균일하게 나타났다(Fig.
4b). 베타카로틴 강 화 콩에서 보이는 농업적 형질의 약세는 모본과의 여교잡으 로 개선될 수 있을 것으로 판단된다.
Fig. 4.Photographs of transgenic plants growing in a greenhouse and transgenic soybean seeds.

도입유전자의 삽입위치 및 도입유전자의 구조 분석
선발된 4계통의 형질전환 콩에 대한 정확한 T-DNA 도입 위치를 확인하기 위해 도입유전자 인접염기서열 분석을 수행 하였다. Adapter PCR을 수행한 결과 분석된 4계통 모두에서 LB 와 RB 방향의 메인 밴드 각각 한 개씩을 얻을 수 있었다 (Fig.
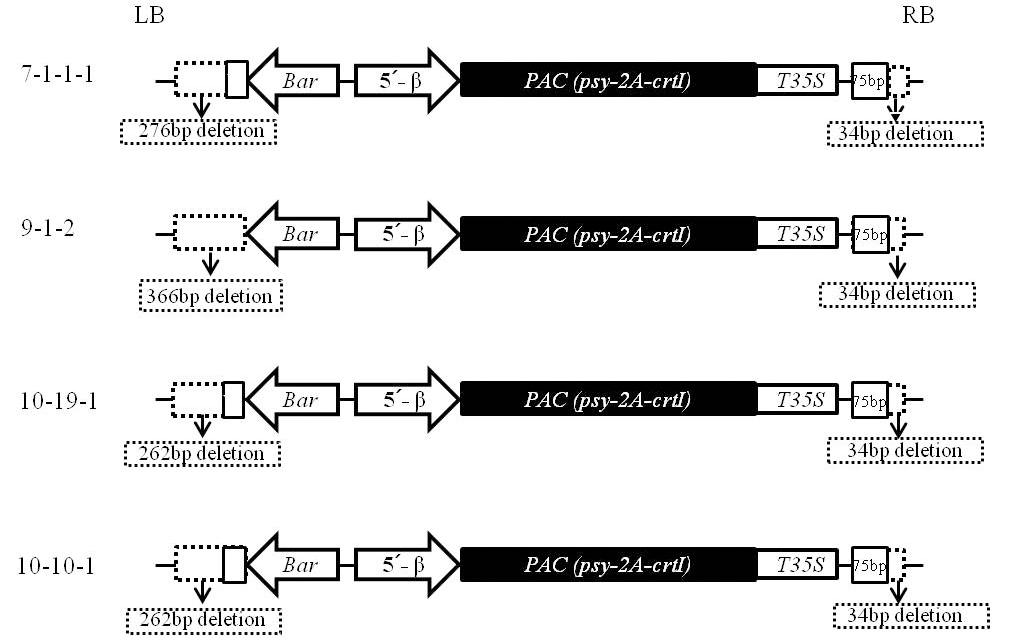
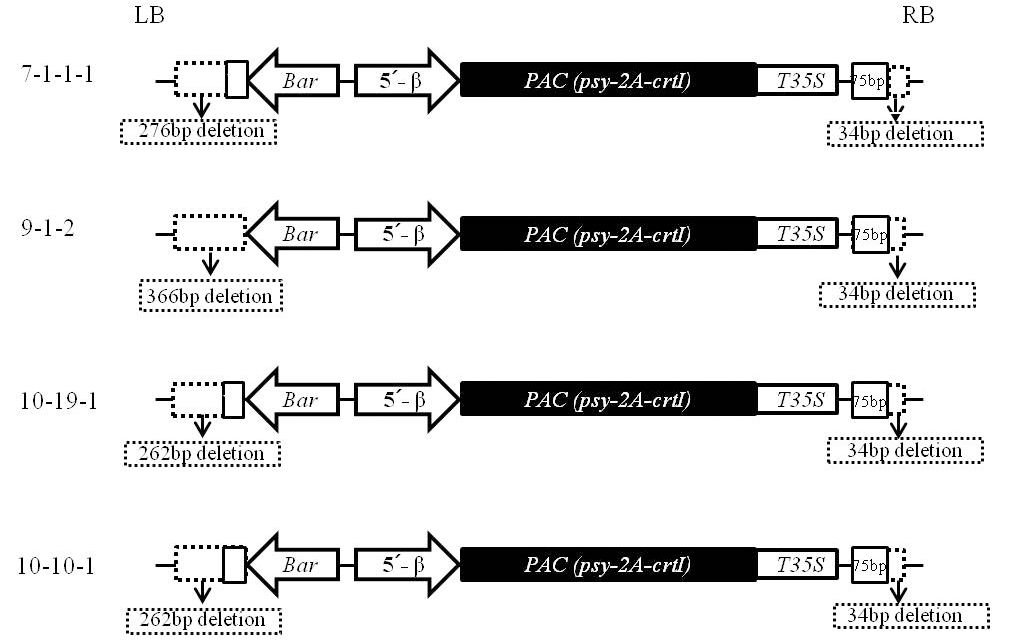
5). PCR 증폭산물에 대한 염기서열을 분석하여 도입유 전자와 연결된 콩의 genomic DNA 분석결과 T
6세대인 7-1-1-1 계통은 14번 염색체의 DnaJ/Hsp40 cysteine-rich domain superfamily protein (Glyma14g12130.1) 유전자의 9.4 kb upstream의 intergenic 부위에 도입유전자가 삽입된 것으로 확인되었다(Table
2). 또한 T
5세대인 9-1-2 계통은 11번 염색 체의 bromo-adjacent homology (BAH) domain-containing protein (Glyma11g02560.1) 유전자의 1 kb upstream의 intergenic 부위에, 10-10-1 계통은 13번 염색체의 Glyma13g20460.1 유전자의 0.237kb upstream intergenic 부위에 도입유전자가 삽입되어 있는 것으로 확인되었다. 그러나, 10-19-1 계통은 13번 염색체의 Glyma13g20460.1 유전자의 exon 부위에 도 입유전자가 삽입되어 있었다. Glyma13g20460 유전자는 leucinerich pentatricopeptide repeat (PPR) protein을 전사하는 부 위로 콩에서의 기능은 알려져 있지 않지만, 애기장대에서의 연구결과 ABA signaling 대사에 관여해 abiotic 또는 biotic 스트레스에 의해 발생되는 활성산소종(reactive oxygen species) 을 조절하는 것으로 알려져 있다(
Laluk et al. 2011).
Fig. 5.The PCR products to left (a) and right (b) border region of T-DNA for flanking sequence analysis. M, molecular marker; W, non-transformed wild-type Kwangan; Lanes 1-4 indicate transgenic soybean line 7-1-1-1, 9-1-2, 10-19-1, and 10-10-1, respectively.
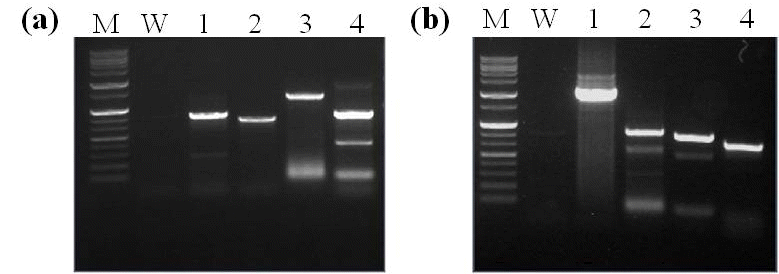
Table 2.Analysis of integration site of T-DNA in the genome of β-carotene enhanced transgenic soybean plant lines.
Table 2.
|
Line |
Chr. |
Insertion site |
Insertion type |
Adjacent Genes |
Generations |
|
|
7-1-1-1 |
14 |
10873131−10872988 |
Intergenic |
Glyma14g12130.1 (9.469kb upstream) |
T6
|
|
9-1-2 |
11 |
1680448−1680251 |
Intergenic |
Glyma11g02560.1 (0.993kb upstream) |
T5
|
|
10-19-1 |
13 |
23919415−23919574 |
Exon |
Glyma13g20460.1 |
T5
|
|
10-10-1 |
13 |
23918985−23919456 |
Intergenic |
Glyma13g20460.1 (0.237kb upstream) |
T5
|
T-DNA 삽입부위에서의 염기서열의 결실, 변형, 재배열 현 상은 여러 식물체에서 분석되었으며 염기서열의 결실은 RB 부 분보다는 LB 부분에서 빈번한 것으로 보고된 바 있다(
Gheysen et al. 1991,
Mayerhofer et al. 1991,
Nacry et al. 1998). T-DNA border 부분에 대한 염기서열 분석결과에서 4계통 모 두에서 정도의 차이는 있지만 DNA 결실이 확인되었다. 구체적 으로 T-DNA의 LB 부위에서는 7-1-1-1 계통은 276 bp, 10-19-1 및 10-10-1 계통은 262 bp, 9-1-2 계통은 13 bp의
nos terminator를 포함하여 366 bp의 DNA 서열의 결실이 발생되 었다. 또한 RB 부위에서는 공통적으로 34 bp의 T-DNA 서열 이 결실된 것으로 확인되었다. RT-PCR 결과(Fig.
3b)로 판단 할 때 9-1-2 계통의 13 bp의 nos terminator 결실은
bar 유전자 의 발현에는 큰 영향을 끼치지는 않는 것으로 사료된다.Fig .
6
Fig. 6.Diagram of T-DNA structure of transgenic soybean lines. The dotted box represent deleted region of T-DNA.
GM작물의 상업화를 위해서는 GMO 안전성평가가 요구된 다. 그러나 식품 및 환경에 대한 영향을 평가하기 위한 GM작 물의 안전성평가는 많은 비용과 전문인력이 필요하다. 따라서 안전성평가 결과를 성공적으로 이끌기 위해서는 분자생물학 적 평가 등을 이용한 안전성평가 재료의 선정이 매우 중요하 다. 본 연구에서는 안전성평가의 재료확보를 위해 multiple copy의 도입유전자가 삽입된 베타카로틴 형질전환 콩의 후대 계통을 육성하여 단일 도입유전자가 콩 염색체의 intergenic 부위에 삽입된 3개의(7-1-1-1, 9-1-2, 10-10-1) 후대계통을 선발하였다. 향후 선발된 계통을 이용하여 복수세대(multigeneration) 에서의 도입유전자의 안정적 유지와 발현 및 전체 T-DNA 구조분석을 수행할 예정이다.
적 요
국내 품종인 광안콩을 이용해 베타카로틴 성분이 강화된 유전자변형 콩이 최근 개발되었다. 본 연구에서는 다수의 도 입유전자가 염색체에 삽입된 유전자변형 콩을 후대 육성하여 유전자분리를 통해 single T-DNA가 도입된 후대계통을 선발 하여 작물재료를 육성하고, 또한 도입유전자 삽입위치를 분석 하여 GMO 안전성평가를 위한 기초자료로 활용하고자 하였 다. 형질전환 콩의 후대 계통에 대한 Southern blot 분석으로 4계통의 단일 도입유전자가 도입된 형질전환 계통을 선발하 였다. 도입위치 분석결과 3계통(7-1-1-1, 9-1-2, 10-10-1)에 도입된 T-DNA는 염색체의 intergenic 부위에 삽입되어 있는 반면, 10-19-1 계통의 T-DNA는 14번 염색체내의 유전자 exon 부위에 삽입된 것으로 분석되었다.
사 사
본 연구는 2015년도 농촌진흥청 국립농업과학원 농업과학 기술 연구개발사업(과제번호: PJ010902) 및 농촌진흥청 차세 대 바이오그린21 박사후연수과정 지원사업에(과제번호: PJ 011188) 의해 수행되었습니다.
References
- 1. Bugert P, Decker S, Klüter H. Improved PCRwalking for large-scale isolation of plant T-DNA borders. Bio. Techniques 2001. 30: 496-504.
- 2. De Buck S, Windels P, De Loose M, Depicker A. Single-copy T-DNAs integrated at different positions in the Arabidopsis genome display uniform and comparable β-glucuronidase accumulation levels. Cell Mol. Life Sci 2004. 61: 2632-2645.
- 3. Doyle JJ, Doyle JL. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin 1987. 19: 11-15.
- 4. Gheysen G, Villarroel R, Van Montagu M. Illegitimate recombination in plants: a model for T-DNA integration. Genes Dev 1991. 5: 287-297.
- 5. Ha SH, Kim JB, Park JS, Lee SW, Cho KJ. A comparison of the carotenoid accumulation in Capsicum varieties that show different ripening colours: Deletion of the capsanthin-capsorubin synthase gene is not a prerequisite for the formation of a yellow pepper. J. Exp. Bot 2007. 58: 3135-3144.
- 6. Ha SH, Liang YS, Jung H, Ahn MJ, Suh SC, Kweon SJ, Kim DH, Kim YM, Kim JK. Application of two bicistronic synstems involving 2A and IRES sequences to the biosynthesis of carotenoids in rice endosperm. Plant Biotech. J 2010. 8: 928-938.
- 7. James C. Global status of commercialized biotech/GM crops: 2013. ISAAA Brief No. 46 2013. ISAAA. Ithaca, NY.
- 8. KBCHDomestic and international trend. Biosafety 2015. 16: 83-89.
- 9. Kim MJ, Kim JK, Kim HJ, Pak JH, Lee JH, Kim DH, Choi HK, Jung HW, Lee JD, Chung YS, Ha SH. Genetic modification of the soybean to enhance the β- carotene through seed-sepcific expression. PLOS One 2012. 7: -e48287.
- 10. Laluk K, Abuqamar S, Mengiste T. The Arabidopsismitochondria-localized pentatricopeptide repeat protein PGN functions in defense against necrotrophic fungi and abiotic stress tolerance. Plant Physiol 2011. 156: 2053-2068.
- 11. Mayerhofer R, Koncz-Kalman Z, Nawrath C, Bakkeren G, Crameri A, Angelis K, Redei GP, Schell J, Hohn B, Koncz C. T-DNA integration: a mode of illegitimate recombination in plants. EMBO J 1991. 10: 697-704.
- 12. Nacry P, Camilleri C, Courtial B, Caboche M, Bouchez D. Major chromosomal rearrangements induced by T-DNA transformation in Arabidopsis. Genetics 1998. 149: 641-50.
- 13. Qin Y, Ahn HI, Kweon SJ, Baek SH, Shin KS, Woo HJ, Cho HS, Lee JH, Lim MH. Molecular characterization of transgenic rice producing resveratrol. Plant Breed. Biotech 2013. 1: 406-415.
- 14. Schubert D, Lechtenberg B, Forsbach A, Gils M, Bahadur S, Schmidt R. Silencing of Arabidopsis T-DNA transformants: the predominant role of gene-specific RNA sensing mechanism versus position effects. Plant Cell 2004. 16: 2561-2572.
- 15. Stahl W, Sies H. Antioxidant activity of carotenoids. Mol. Aspects. Med 2003. 24: 345-351.
- 16. Ye X, Al-Babili S, Kloti A, Zhang J, Lucca P, Beyer P, Potrykus I. Engineering the provitamin A (β- carotene) biosynthetic pathway. Science 2000. 287: 303-305.