적요
Fusarium wilt, caused by a fungal pathogen Fusarium oxysporum f. sp. conglutinans (Foc), is a destructive disease of Brassica oleracea var. capitata. Currently, there are reports concerning molecular markers linked to Fusarium wilt disease resistance gene (FocBo1) in cabbage. In this study, we have studied a relationship between the FocBo1 gene and six cabbage breeding lines obtained from Asia Seed Company. The six cabbage breeding lines were evaluated resistance levels to Foc by combing of disease phenotypes and fungal biomass in planta. Of 6 breeding lines, KR-518 and OK-517 presented strong resistance and RK-P6-1, moderately resistance, whereas MT-624, JK-2, and HY-164, were susceptible to Foc. To define the correlation between the wilt disease resistance phenotype and the presence of resistance FocBo1 gene, we employed RT-PCR with MTK-C marker, which is known to be linked to FocBo1 resistant gene. The results exhibited inaccuracy in differentiating resistant and susceptible plants with genomic DNA samples, but it shows 100% correlation with cDNAs, indicating there is exceptional breeding line that contains FocBo1 gene in its genome with no expression. After cloning and sequencing the MTK-C product, a specific molecular marker, FocBo1-C, was developed which amplify a single, easily resolved band from cDNA of cabbage breeding lines with FocBo1 gene. Altogether, these results indicate that the three cabbage breeding lines in resistant category contain the resistance FocBo1 gene and moreover, the expression level of FocBo1 affect the degree of resistance of cabbage breeding lines, suggest the semidominant nature of FocBo1 gene in cabbage.
서 언
양배추(Brassica oleracea var. capitata)는 십자화과(Brassica Family)에 속하는 채소 작물로써, 한국 내 각 지역에 알맞은 재배 작형이 분화되어 있어 주년 재배가 가능하며, 국내에서 는 배추, 고추, 무와 같은 다른 십자과 작물들에 비해 양배추 의 소비량이 낮지만, 음식의 다양화 및 건강에 대한 관심으로 소비량 및 생산량이 점차 증가하고 있는 추세이다.
최근 양배추에 대한 병해로 곰팡이에 의한 시들음병이 발 생하여, 전 세계 양배추 생산량 감소의 큰 원인으로 지목 되 고 있다(
Brown & Proctor 2013). 시들음병을 일으키는 원인 균은
Fusarium oxysporum으로 알려져 있으며, 이 균은 120 종이 넘는 분화형이 존재하는 다양한 변이를 가진 종으로, 각 각의 분화형은 다양한 식물군에 걸쳐 특정 기주에 병을 일으 키는 것으로 알려져 있다(
Michielse & Rep 2009). 양배추에 병 을 일으키는 분화형은
Fusarium oxysporum f. sp.
conglutinans (
Foc)이며 무를 제외한 다른 배추과 작물에도 병을 일으키고 있는 것으로 알려져 있다. 이 균에 의한 주요 병징은 잎의 황 화, 물관의 변색, 낙엽 등으로, 시일이 지나면 감염된 식물은 결국 고사하게 된다(
Michielse & Rep 2009,
Moon et al. 2001).
Foc는 토양에서 수십 년 이상 생존이 가능한 토양전 염성 병원균으로, 이의 방제를 위해 배추과 이외의 작물과의 윤작, 종자 소독 후 파종, 토양 살균제의 사용, 석회 사용 및 질소비료의 과용 금지 등의 방법들이 적용되고 있으나, 이러 한 방법들은 재배 토양에서 이 병균을 완전히 제거할 수 없을 뿐 아니라, 환경오염 등의 문제가 생기기 때문에, 시들음병 저 항성 품종의 사용이 가장 효과적인 방제 방안으로 제시되어 지고 있다(
Baik et al. 2011,
Lee et al. 2014).
미국에서는 20세기 초부터 시들음병에 저항성을 갖는 양배 추를 육종하기 위한 연구가 시작되었다(
Walker & Wellman 1927). 시들음병에 대한 양배추 저항성 반응은 Type A와 Type B, 두 가지 유형으로 크게 나누고 있다. Type A 저항성 은 단인자 우성 유전을 하며, 토양 온도가 24°C 이상으로 상 승하여도 시들음병에 대한 저항성 표현형이 안정적으로 나타 나는 것으로 알려져 있는 반면, Type B 저항성은 다양한 유 전 인자에 영향을 받으며 토양 온도가 24°C 이상으로 상승하 면 저항성이 불안정 해진다는 특성을 가지고 있다(
Blank 1937). 따라서 온도 변화에도 안정적이며, 단일 우성 유전자 에 의해 저항성이 조절되므로, 양배추 육종 시에 병저항성을 쉽게 획득시킬 수 있다는 장점 때문에 Type A 저항성 양배추 품종이 상업적으로 많이 이용되고 있다(
Smith 1930,
Blank 1937,
Walker 1953). 그러나 Type A 저항성 유전자를 이용 해 시들음병 저항성 양배추를 육종한지 80년 이상이 되었음 에도 Type A 유전자를 실제로 클로닝 하기 위한 시도는 근래 에 들어서야 이루어졌으며, 최근 양배추에서 Type A 저항성 에 의해 시들음병을 조절하는 것으로 여겨지는
FocBo1 라는 저항성 유전인자에 대한 연구 내용이 발표되었다(
Pu et al. 2012,
Shimizu et al. 2015).
Pu et al. (2012)은 시들음병 저 항성 양배추와 감수성 브로콜리를 교잡, 이들의 자손 세대에 서 시들음병에 대한 표현형을 관찰하여, 시들음병 저항성을 조절할 것으로 예상되는 단일 우성 대립 유전자를 발견하고 이를
FocBo1이라 명명하였다(
Pu et al. 2012). 그리고
FocBo1유전자를 토대로 시들음병 저항성 양배추를 구분 할 수 있는 분자 마커인 MTK-C에 대한 연구결과를 발표하였다. MTK-C를 이용하여 일본에서 상업적으로 이용되고 있는 양 배추 품종의 genomic DNA로부터
FocBo1 유전자의 존재를 확인 한 결과, 모든 저항성 품종에서는 MTK-C에 의해 증푹 된 DNA 조각이 확인 되지만 감수성에서는 확인되지 않았다 (
Shimizu et al. 2015).
본 연구에서는 국내에서 보유한 시들음병관련 양배추 육종 계통의 시들음병 처리에 대한 반응을 병징과 식물 생체 내 곰 팡이 바이오 매스 정량을 통하여 정확히 진단하고, 또한, 이들 계통의 병관련 반응을 밝혀진 FocBo1저항성 유전자와의 상 관성과 함께 MTK-C마커를 이용하여 FocBo1 유전자의 존재 유무 및 발현을 연구 하였다.
재료 및 방법
식물 재배
본 연구를 위하여, 아시아 종묘회사로부터 7가지 양배추 품종 종자들을 분양받았다. 종자들을 순차적으로 70% 에탄 올 용액에서 1분간 1회, 12% 차아염소산나트륨용액에 15분 간 1회 소독하고, 멸균수에 5회 세정하였다. 4 × 8 육묘용 연 결 포트(원농사)에 멸균된 원예용 상토2호(원농사)를 넣고 각 소독된 종자들을 16 포트씩 그리고 포트 당 2립씩 파종하여, 4°C에서 4일간 저온처리 후 식물 생장실(23°C)에서 재배하 였다. 종자가 발아한 후 생장이 고른 개체로 포트 당 1주씩 남겼고, 파종 후 17일 재배한 유묘를 실험에 사용하였다.
시들음병 접종원 준비 및 접종
한국화학연구원으로부터 분양받은
Fusarium oxysporum f. sp.
conglutinans균주를 potato dextrose agar(Becton, Dickinson and Co., USA) 배지에 접종하고, 22°C에서 7일 동안 배양한 균총으로부터 균사조각을 떼어내어 malt extract broth (Becton, Dickinson and Co., USA) 배지에 접종하여 28°C, 암상태에 서 180rpm으로 4일 동안 진탕배양하였다. 배양한 균주를 8겹 거즈로 걸러 균사를 제거한 후, 수확된 포자를 광학현미경 하 에서 hemocytometer로 측정하고 물로 희석하여 1.0 × 10
7 conidia/ml의 포자 농도로 준비하였다(
Baik et al. 2011). 온 실에서 재배한 유묘의 뿌리를 수돗물로 세척하여 흙을 제거 하고 준비된 포자 현탁액에 30초간 침지하여 접종한 후, 4 × 8 육묘용 연결 포트(원농사)에 멸균한 원예용 상토 2호(원농 사)를 넣고 접종한 유묘를 이식하였다(
Song et al. 1996). Mock 처리에는 동일한 방법으로 유묘 뿌리를 수돗물에 침지 하여 멸균된 상토에 이식하였다.
시들음병원균이 접종된 유묘들이 이식된 포트들을 48 cm × 23 cm × 15 cm(H) 크기의 육묘용 투명 플라스틱 덮개로 덮어 습도를 유지하였고, 하루에 16시간 동안 광을 조사하면 서 재배하였다. 접종 19일 후에 식물체 잎의 황화 및 식물체 생장 정도에 따른 발병도를 조사하였다(
Thatcher et al. 2009). 시들음병의 병조사 기준은 0 = 병증 없음, 1 = 식물체의 황 화, 2 = 고사 등 의 3단계로 하였다(
Shimizu et al. 2014). 시 들음병의 평균 발병도가 1.0이하인 경우에는 저항성, 1.0 – 1.85은 중도저항성 (MR), 1.86 이상은 감수성으로 판정하였 다. 모든 실험은 각 육종계통 별로 8~12 이상의 개체를 가지 고 수행하였으며, 독립적인 실험으로 2회 실시하였다.
시들음병균 접종 19일 된 식물체를 샘플링하여 genomic DNA (gDNA)를 DNeasy Plant Minikit (Qiagen, USA)을 이용하여 추출하였고, 실험 방법은 제조사의 메뉴얼을 따라 수행하였다. 양배추 및
Foc의 특이 유전자 발현 정도를 측정 하기 위해 CFX connect
TMReal Time PCR system (Bio-Rad, USA)을 이용하여 quantitative realtime-PCR (qRT-PCR)을 수행하였다. 시들음병균의 생물량 측정을 위하여
Foc의
glyceraldehyde 3-phosphate dehydrogenase (
GPD) 유전자 특이적 인 프라이머 쌍(Table
1)을 사용하여 다음과 같은 과정으로 Q-RT-PCR를 수행하였다(
Thatcher et al. 2009). PCR반응 용액은 5 μl 2x iQ
TMSYBR® Green Super Mix (Bio-Rad, USA), 1 μl
GPD 프라이머쌍(10 pmol, Bioneer, Korea), 3 μl HPLC-water, 그리고 10-15ng gDNA 주형을 혼합하여 총 10 μl로 조성하였다. Q-RT-PCR은 95°C, 3분 denaturation 1 회, 95°C, 20초 denaturation – 60°C, 20초 annealing – 72°C, 20초 extension을 40회 반복, 95°C, 10초간 반응시킨 후, 65°C에서부터 95°C까지 5초간 0.5°C씩 상승시켜 melt curve analysis를 실행하였다.
B.oleracea의
Actin (
Act1)유전 자 특이 프라이머 쌍(Table
1)을 이용하여 위와 같은 방법으로 Q-RT-PCR을 수행하고,
Foc GPD 측정값을 평준화하였다.
Table 1.Primers used in this study.
Table 1.
|
Primers |
Sequence (5’ 3’) |
Reference |
Application |
|
|
F. oxysporum GPD_F
|
AAGGGTGCTTCTTACGACCA |
Thatcher et al.2009
|
Q-RT-PCR |
|
F. oxysporum GPD_R
|
ATCGGAGGAGACAACATCGT |
Thatcher et al.2009
|
Q-RT-PCR |
|
MTK-C-F |
GAGCTCTCTTTCTCTACAAGTCAAGG |
Shimizu et al.2015
|
RT-PCR |
|
MTK-C-R |
GTTAGCTGAGTAACGAGATAGACCATG |
Shimizu et al.2015
|
RT-PCR |
|
BoActin_F
|
CAGTGTCTGGATCGGTGGTT |
this study |
RT-PCR |
|
BoActin_R
|
GAGGCATACATCAATTCGATCAC |
this study |
RT-PCR |
|
BoActin_QF
|
CAGTGTCTGGATCGGTGGTTCC |
this study |
Q-RT-PCR |
|
BoActin_QR
|
AAACCCCAGCTTTGAGCCTTCT |
this study |
Q-RT-PCR |
|
FocBo1-C_F
|
AATATTGCATACGAGTGTGTCTTCGC |
this study |
RT-PCR |
|
FocBo1-C_R
|
TGCATTGGGGTATGGTGGTACTTTAG |
this study |
RT-PCR |
분자 마커 검정을 위한 reverse transcription PCR (RT-PCR) 증폭 및 sequencing
시들음병균 접종 8일 된 유묘의 전체 RNA를 RNAiso Plus (Takara, Japan)을 이용하여 분리하였고, 실험은 제조사의 매 뉴얼에 따라 수행하였다. 추출한 전체 RNA 1
μg, oligodT (Bioneer, Korea) 2 μl와 HPLC-water를 혼합하여 총 부피가 11 μl되도록 한 후, 65°C 항온수조에서 10분간 반응시켰다. 반응시킨 RNA와 5×First Strand Buffer (Invitrogen, USA) 5 μl, 2.5 mM dNTP 6 μl, Moloney Murine Leukemia Virus (M-MLV) reverse transcriptase (Invitrogen, USA) 1 μl, Solg
TMRNase Inhibitor (SolGent, Korea) 1 μl, 0.1 M DTT (Invitrogen, USA) 1 μl를 혼합하여 37°C 항온수조에 서 1시간, 65°C 항온수조에서 10분간 반응하여 cDNA를 합 성하였다. 합성된 cDNA,
FocBo1-C Marker 염기서열 특이 프라이머 쌍(Table
1), Accupower® PCR Premix (Bioneer, Korea)를 혼합하여 다음과 같은 과정으로 MJ research PTC-200 thermocycler (Bio-Rad, USA)을 이용하여 RT-PCR 수행하였다. 95°C에서 2분 denaturation 1회 후, 95°C, 30초, denaturation - 59°C, 30초, annealing – 72°C, 1분 extension을 29회 반복하고 72°C에서 5분간 반응하였다.
B. oleracea의
Actin 유전자 특이 프라이머 쌍을 이용하여 다 음과 같이 유전자를 증폭하였다. 95°C에서 2분 denaturation 1회 후, 95°C, 30초, denaturation – 55°C, 30초, annealing – 72°C, 30초 extension을 25회 반복하고 72°C에서 5분간 반응하였다. Genomic DNA의 유전자 증폭은 26회 반복을 제외하고 위와 동일한 조건으로 PCR이 수행되었다. PCR증 폭이 끝난 생산물은 EtBr(0.5
μg/ml)이 함유된 1.5% 아가로스 젤에서 분리 후, UV 아래에서 확인하였다. 확인된 증폭 산물들 을 Accuprep® PCR Purification kit (Bioneer, Korea)을 이 용하여 정제한 후, T&A Cloning kit (RBC, Taiwan)의 T-vector내로 클로닝하고 Ampicillin (0.1 mg/ml)이 함유된 Luria Bertani (LB)고체배지에서 콜로니를 선별하였다. 선별된 콜로니 배양액에서 Accuprep® Nano-plus Plasmid mini extraction kit (Bioneer, Korea)을 이용하여 플라스미드DNA 를 분리하고 염기서열 분석을 의뢰하였다.
결과 및 고찰
양배추 품종의 시들음병 저항성 조사
본 실험에서는 아시아종묘회사로부터 분양 받은, 시들음병 관련 6가지 양배추 육종계통들의 시들음병에 대한 반응성을 실험실 환경과 분자생물학적 기법을 이용하여 정확히 평가하 고자 하였다. 우선, 시들음병에 대한 저항성 표현형의 대조구 로써 상업적으로 판매가 되고 있는 ‘대박나’(아시아 종묘)를 이용하였다(
Baik et al. 2011). 시들음병균의 포자 (1.0 × 10
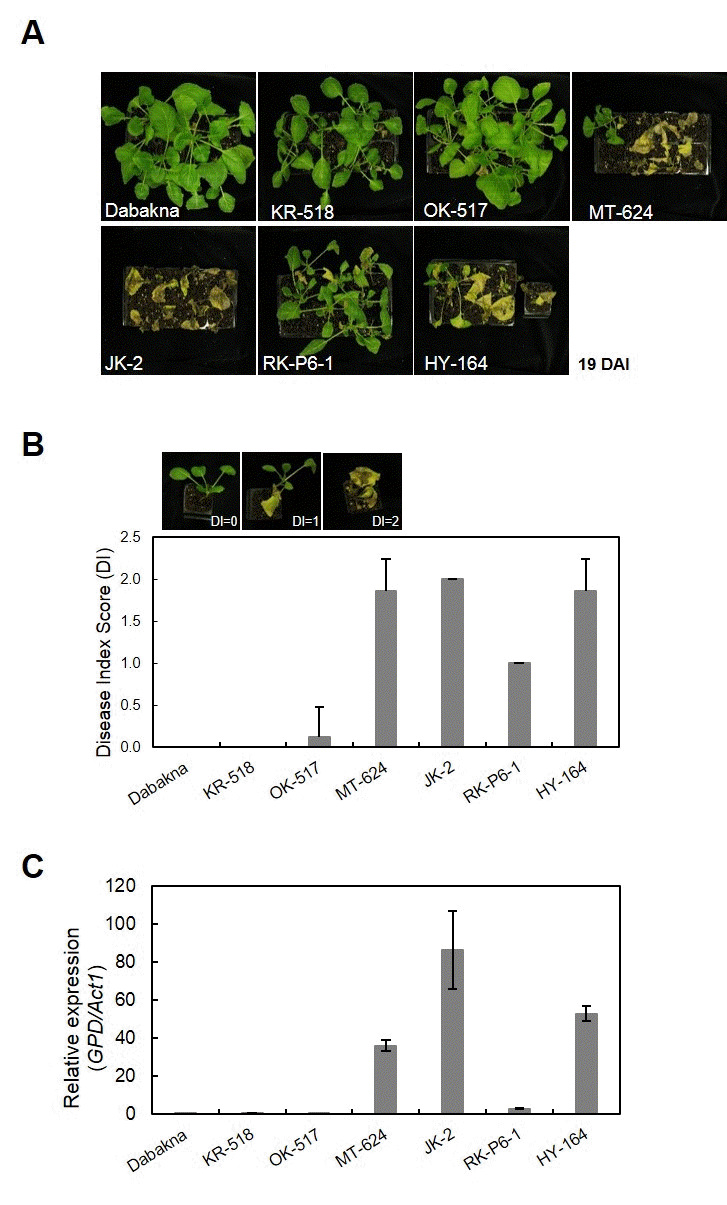
7 conidia/ml)를 준비된 식물개체에 처리하고, 병 접종 19일 후 상층부의 병징을 관찰하였다(Fig.
1A 및 Table
2). 조사한 6 가지 계통 중 KR-518, OK-517은 두번의 실험에서 모두 발 병도 (Disease index, DI)가 각각 0.00, 0.13으로 강한 저항성 으로 나타났고, RK-P6-1은 DI가 1.00으로 중도 저항성 표현 형을 보였으며, JK-2, HY-164는 1.86 이상의 높은 발병도를 나타내어 시들음병에 대해 감수성임을 확인하였다(Fig.
1 및 Table
2). 저항성 계통 중 KR-518은 식물 전체에서 아무런 병징을 보이지 않은 반면, OK-517의 경우에는 10% 정도의 개체가 떡잎이 황화되는 병징을 보였다. 감수성 계통인 JK-2 와 HY-164의 경우에도 병징의 발현 속도에서 차이를 보였는 데, JK-2가 HY-164에 비해 식물이 4-5일 정도 더 빨리 괴사 되는 특징을 보였다(data not shown).
Fig. 1.Development of Fusarium wilt disease in cabbage breeding lines. (A) Seventeen days old cabbage seedlings were inoculated by dipping the root tips of seedlings in spore suspensions of Foc (inoculum density of 1.0 × 107 spores/ml). Disease symptoms were monitored over a period of 21 days. Photos were taken 19 days after infection (DAI). (B) Disease symptoms were scored at 19 DAI from 0 to 2 as follows: DI (disease index); 0, no symptoms or chlorosis on the cotyledons; 1, necrotic or yellowish symptoms in half of plants; and 2, necrotic or water-soaked symptoms in all plants. R, resistant (DI ≤ 0.125); MR, moderate resistant (1.00 ≤ DI < 1.85); S, susceptible (DI ≥ 1.86). Each value represents the mean disease index score of two runs with 7~12 replicates each. (C) Foc biomass in cabbage breeding lines. Values are relative to B. oleracea DNA by assaying fungal GPD and B. oleracea iASK sequences from the leaves of inoculated plants by quantitative real-time PCR. Error bars indicate standard error of three technical repeats of one biological batch.
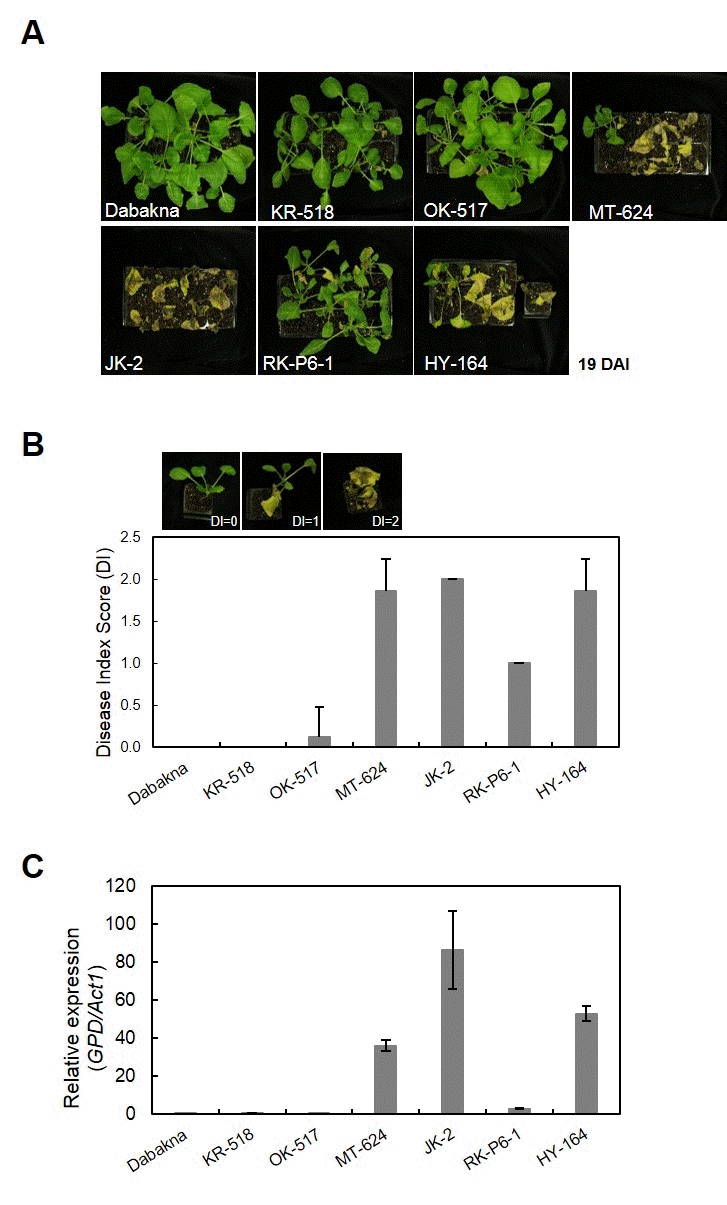
Table 2.Summary of characteristics of six Brassica oleracea var. capitata breeding lines.
Table 2.
|
Cultivar |
Lines |
Head Shape |
Foc wilt disease phenotypesz
|
|
|
Dabakna |
F1 |
Flat |
R |
|
KR |
518 |
Flat |
R |
|
OK |
517 |
Flat |
R |
|
MT |
624 |
Round |
MS |
|
JK |
JK-2 |
Round |
S |
|
RK |
P6-1 |
Oval |
MR |
|
HY |
164 |
Flat |
S |
MT-624는 DI가 1.89로 높은 감수성을 나타내었으나, 2회 의 반복 실험을 통해 본 결과 병징이 처리 환경에 따라 달라 지는 것을 볼 수 있었다. Baik et al. (2011)의 연구에서 양배 추 뿌리를 포자 현탁액에 담근 즉시 꺼낸 경우 감수성 품종에 서도 시들음병의 발병도가 낮아진다는 보고가 있으며(
Baik et al. 2011), MT-624의 경우도 포자의 상태 또는 균의 처리 방 법에 따라 민감하게 반응하는 품종으로 생각된다.
위 실험에서 MT-624와 RK-P6-1 경우 반복 실험에서 병 징 차이만으로 시들음병의 저항성 정도를 판단 하기에 어려 움이 있으므로, 이를 보완하기 위해 식물체내
Foc의 생체질 량을 측정하고자 하였다. 최근
Li et al. (2015)의 실험에서 저항성과 감수성 양배추에 시들음병균을 처리하였을 때 감수 성 품종은 병처리 후 7일째부터 시들음병균이 양배추의 뿌리 에서부터 상층부의 잎과 줄기 부분까지 증식 하는 것을 확인 한 반면에 저항성 품종에서는 상층부에서 시들음병균이 전혀 확인되지 않는다는 보고가 있었다(
Li et al. 2015). 더불어,
Thatcher et al. (2009)은 시들음병균을 처리한 애기장대 잎 에서 식물체에서는 존재하지 않는 곰팡이 유전자 특이 프라 이머를 이용하여 식물체내 곰팡이의 생체질량을 측정한 바 있다(
Thatcher et al. 2009). 이들은 이 실험결과로부터, 시들 음병균에 의한 식물 기주의 괴사가 많이 진행되었을 경우 저 항성과 감수성 식물의 잎 조직으로 확인되는 곰팡이의 생체 질량 수준이 현저히 다르게 나타나는 것을 확인하였다. 육안 으로 판별하는 병징조사의 경우 실험자에 의한 오류 발생 가 능성을 배제할 수 없다(
Trusov et al. 2013). 따라서 본 실험 에서는
Foc의 genomicDNA 내 존재하는
glyceraldehyde 3-phosphate dehydrogenase (
GPD) 유전자의 양과 식물체의 genomic DNA에 존재하는
Actin 유전자의 양을 비교하는 상 대 측정을 통해
Foc가 감염된 식물의 병에 대한 저항성/감수 성을 판단해보고자 하였다. 이를 위해 병처리 후 19일 된 양 배추 식물의 잎에서 genomic DNA를 추출하여 곰팡이
GPD, 양배추
actin 특이프라이머를 이용해 qRT-PCR을 수행하였 다(Table
1 및 Fig.
1C). 저항성 대조구인 대박나 (0.01)와 비 슷하게, OK-517, KR-518 계통의 경우 균의
Foc 병원균의 생체량이 각각 0.55, 0.06로 낮았고 감수성 품종인 JK-2와 HY-164의 경우 86.28과 52.95로 매우 높아 시들음병 저항성 표현형 검정 결과와 일치하는 것으로 나타났다(Fig.
1B,
C). 반면, MT-624는 생물검정에 따른 발병도가 1.89로 높은 감 수성 표현형을 나타내는 계통이나 병원균의 생체량은 36.06 으로 나타났으며, RK-P6-1은 발병도가 1.00인 중도저항성인 데 반해 병원균의 생체량은 3.03을 나타내었다.
이러한 결과는 MT-624과 RK-P6-1의 경우 환경에 의해 조절되어지는 다형적 병 감수성 또는 저항성 메커니즘을 각 각 가지고 있는 계통으로 생각되어진다.
Smith (1930)는 시들음병에 동형접합성 저항성을 가진 양 배추와 동형접합성 감수성 양배추를 교잡시킨 이형접합성 양 배추의 경우, 24°C에서는 시들음병에 대한 저항성을 나타내 나, 26°C이상의 고온에서는 성장이 지연되고 잎이 황화되는 병징이 나타나며 감수성이 증가한다는 연구결과를 발표한 바 있다(
Smith 1930). 또한 저항성 양배추의 F3 line에서 저항성 유전자에 이형접합성을 가진 식물이 시들음병에 대해 완전한 저항성을 가지지 못함을 관찰하였고, 따라서 양배추가 병 저 항성 대립유전자를 하나의 copy 만 가질 경우 시들음병에 대 해 부분적인 저항성을 가지며, 고온에서 시들음병에 대한 감 수성이 높아진다고 하였다(
Pu et al. 2012). 따라서 본 연구에 서 RK-P6-1이 보인 중도 저항성 표현형은 RK-P6-1이 시들 음병 저항성 유전자에 대해 이형접합성을 가짐으로써 나타나 는 결과 일수도 있을 것으로 생각된다.
본 연구에서 국내 보유 6가지 양배추 육종 계통의 시들음 병에 대한 표현형 검정 결과, OK-517과 KR-518은 환경에 의한 변이가 없는 안정적이고 강한 저항성을 가지고 있으며 JK-2와 HY-164은 감수성을 가진 계통으로 나타났다(Table
2). 이는 향후 양배추 시들음병 저항성 품종 육종을 위한 유 전자원으로 유용하게 이용될 수 있을 것으로 생각 된다.
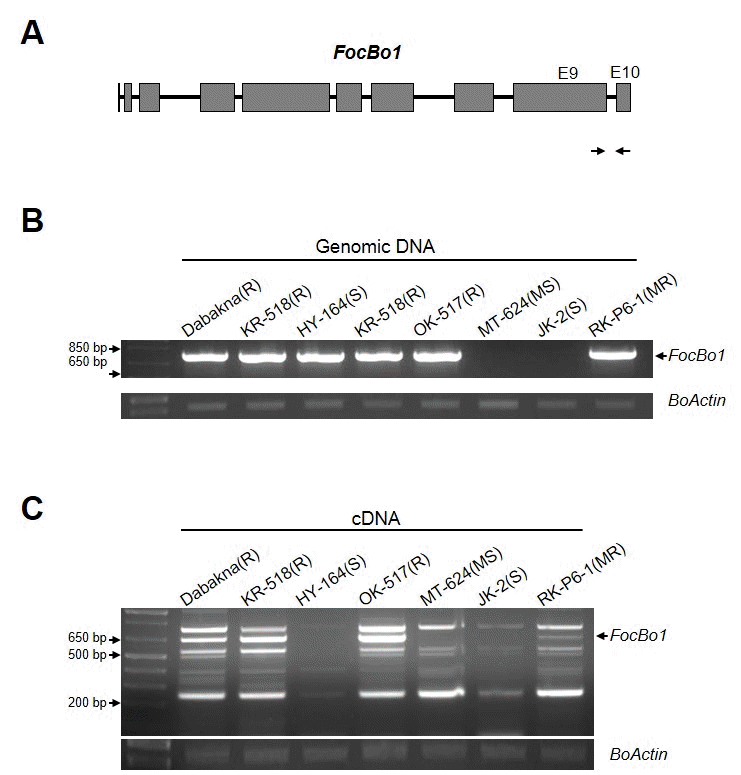
최근
Shimizu et al. (2015)은 시들음병에 대한 표현형과
FocBo1 유전자와의 연관성을 확인 할 수 있는 MTK-C marker를 발표하였는데, MTK-C를 이용하여 증폭되어지는 부분은
FocBo1 유전자의 9번째 exon에서 3’ untranslated region인 것으로 알려져 있다(Fig.
2A)(
Shimizu et al. 2015). 따라서, 본 연구에서는 MTK-C를 이용하여 6가지 육종 계통 의 시들음병에 대한 저항성 또는 감수성 표현형이
FocBo1 유전자와 연관된 것인지 확인하고자 하였다.
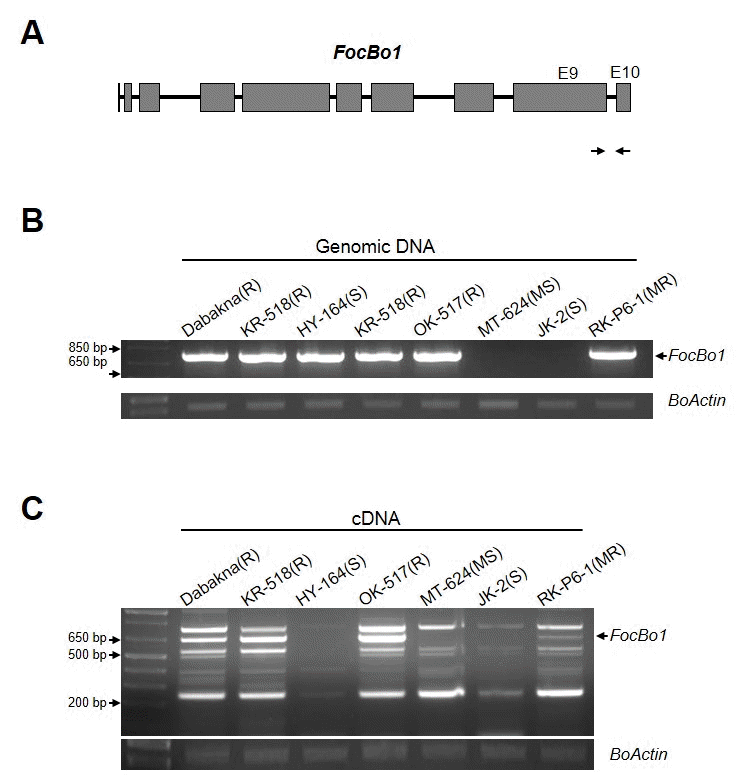
우선,
FocBo1 저항성 유전자의 존재 유무를 확인하기 위 해, 대박나를 포함한 6개 육종 계통의 genomic DNA를 추출 하였다. 이를 주형으로 MTK-C 프라이머 쌍을 이용하여 PCR 를 수행하였다(Fig.
2B). 저항성 품종 대조군인 ‘대박나’ 와 같이 저항성 품종 OK-517, KR-518, 그리고 중도 저항성인 RK-P6-1 품종에서 MTK-C 특이 밴드가 증폭되었으며, 감수 성인 MT-624, JK-2에서는 밴드가 없음을 확인하였다. 하지 만, 기대와 달리 완전 감수성 품종인 HY-164에서도 MTK-C 밴드가 확인되었다(Fig.
2B). Genomic DNA에서는 존재 하 지만, 실제적인
FocBo1 유전자의 발현이 같이하고 있는지 확 인하기 위하여, 각 식물체에서 total RNA를 분리하고, cDNA 를 합성하여,
FocBo1 유전자의 발현을 RT-PCR을 통해 확인 하였다(Fig.
2C). 그림에서 본 바와 같이 다수의 비특이적인 밴드들이 형성되어,
FocBo1 유전자로 예상되는 크기인 600 bp~800 bp사이의 세개의 밴드를 클로닝하여 염기서열을 확 인한 결과, 658 bp 크기의 밴드가 MTK-C 마커에 의해 증폭 된
FocBo1 유전자 산물임을 확인하였다(Fig.
2C). 이 크기의 밴드가 시들음병 저항성 계통인 대박나, KR-518, OK-517과 중도저항성인 RK-P6-1에서만 증폭되고, 감수성을 나타내는 MT-624, JK-2 뿐만 아니라 gDNA에서 저항성 밴드 패턴을 나타냈던 HY-164 계통에서는 증폭되지 않음을 확인하였다 (Fig.
2C). 또한 발현량을 비교 하였을 때, 중도 저항성을 보 이는 RK-P6-1은 저항성 계통에 비해 그 발현량이 적음을 알 수 있었다. 이러한 결과는 국내에서 보유하고 있는 시들음병 저항성 육종 계통들의 저항성 반응이
FocBo1 유전자를 가지 고 있는 것과 함께, 그 발현량이 병 저항성 표현형을 결정하 는 중요한 부분 인 것을 확인 할 수 있었다.
Fig. 2.Classification of the six cabbage breeding lines with MTK-C marker. (A) Schematic diagram of FocBo1 gene structure. Arrows correspond the sequences of MTK-C primers pair. Gray boxes represent exon regions. (B) PCR with the MTK-C marker to verify the present of FocBo1 gene in six breeding lines. Total genomic DNA (gDNA) was extracted from the leaves of each cabbage plants. The amount of BoActin was used as internal control. (C) Expression of FocBo1 gene in six cabbage breeding lines. Total RNA was extracted from each Foc-infected cabbage plants at 8 dpi. Arrows indicates the FocBo1 specific bands. The amount of Boactin transcripts was used as internal control.
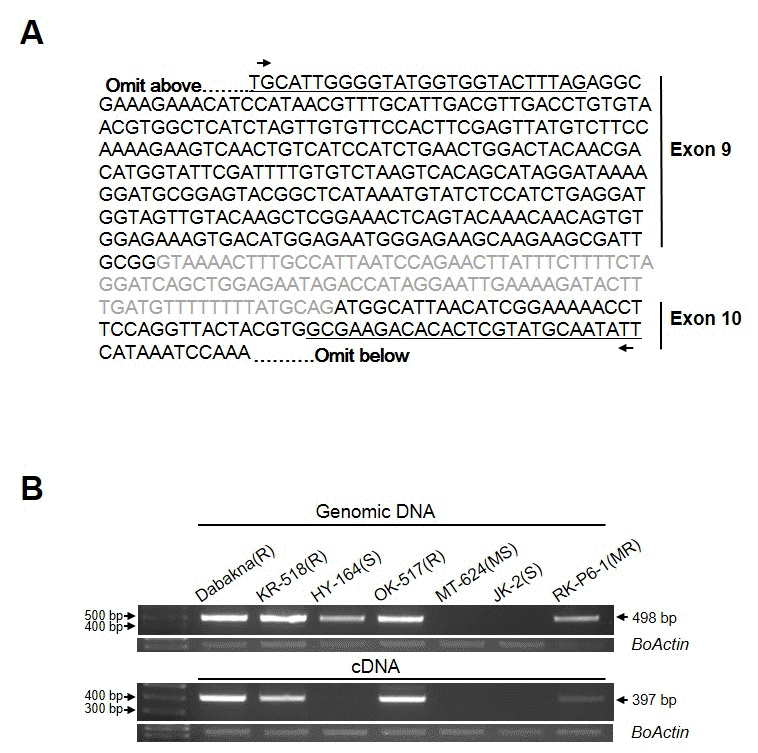
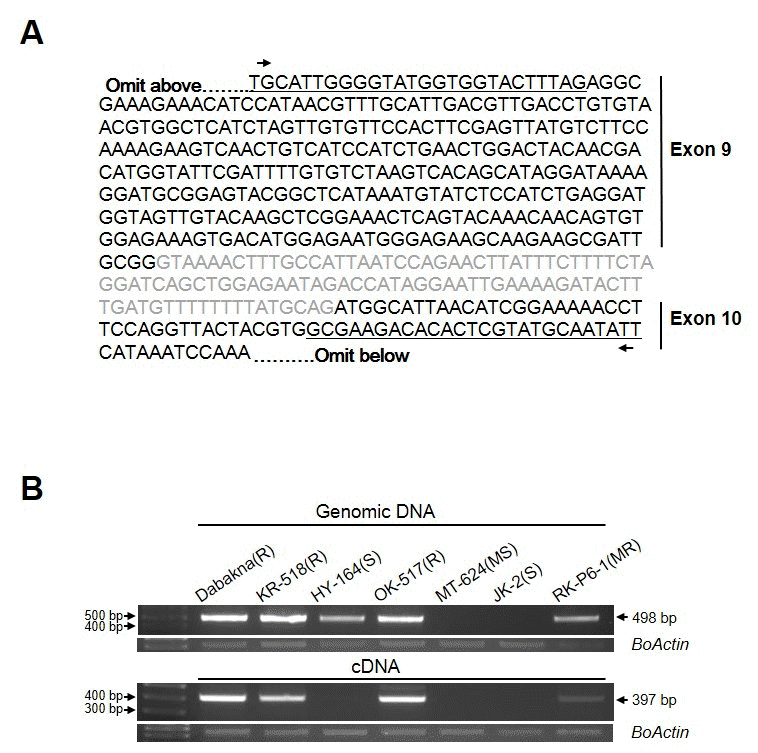
HY-164의 경우 gDNA 상에서는
FocBo1이 존재 하나, 그 발현은 없는 것으로 확인되어 MTK-C에 의해 증폭된 HY-164 genomic DNA 상의 염기서열이 저항성 계통과의 차이가 존 재하는지 확인하기 위해, KR-518과 HY-164 genomic DNA 에서 증폭된 밴드를 클로닝 후 sequencing 하여 염기서열을 확인하였다. 그 결과 염기 서열에서는 두 계통간의 차이가 없 는 것으로 확인 되었다(Fig.
3A). 일본의 ‘RaKuen’ 품종의 경우 gDNA에서 MTK-C 마커 검증 결과 저항성과 동일한 밴드 패턴을 보였으나 표현형은 감수성이었다. 이 품종의 경 우
FocBo1 유전자의 4번째 exon에 mutation이 일어나 저항 성 유전자를 가지고 있으나 정상적인 기능을 하지 못하는 것 으로 알려져 있는데(
Shimizu et al. 2015), HY-164의 경우도 같은 경우가 아닐까 생각 된다. 이의 확인을 위해 HY-164의
FocBo1 유전자의 4번 exon 부분을 클로닝하여 염기 서열을 확인하고, 이에 따른 마커를 개발하는 추가실험이 필요할 것 으로 생각된다.
한편, MTK-C 프라이머 쌍을 이용하여 cDNA를 주형으로 RT-PCR을 수행 하였을 때 여러 크기의 비 특이적인 PCR 산 물들이 확인됨으로(Fig.
2C), 저항성 품종인 KR-518 품종에 서 MTK-C 마커에 의해 증폭되어 얻은 염기서열을 바탕으로 RTPCR 수행 시 단일 밴드를 증폭할 수 있는 새로운
FocBo1-C 프라이머 쌍을 디자인하였다(Fig.
3A및 Table
1). 새 프라이 머 쌍을 이용하여 6가지 육종 계통의 gDNA 및 cDNA에서 PCR을 수행하였을 때 약 400 bp 크기의
FocBo1-C 프라이 머 특이적인 단일 밴드를 확인할 수 있었다(Fig.
3B).
Fig. 3.Analysis of expression FocBo1 with FocBo1-C marker in cabbage breeding lines. (A) Genomic DNA sequence of FocBo1 gene amplified from KR-518. The letters in black indicate exon 9 and 10 regions, respectively, and the sequences of FocBo1-C primers are underlined. (B) Application of FocBo1-C marker on seven inbred lines. Genomic DNA and RNA are extracted from the F. oxysporum inoculated plants after 8 days infection. Arrows indicate PCR products amplified by FocBo1-C primers with a size of 498 bp and 397 bp with genomic DNA or cDNA as template, respectively. The amount of BoActin transcripts were used as internal control.
이상의 결과로부터 KR-518, OK-517이 시들음병 저항성 양배추 육종 자원으로서 활용 가능성을 확인하였고, 국내의 양배추 육종 자원들의 시들음병 저항성이 FocBo1 유전자와 연관되어 있으며, 완전한 저항성을 얻기 위해서는 FocBo1 유 전자의 copy 수도 중요하기 때문에 분자 육종을 위해서는 genomic DNA 수준과 cDNA수준에서의 판별이 모두 필요하 다는 것을 알 수 있었다.
적 요
시들음병은 진균성 병원균인 Fusarium oxysporum f. sp. conglutinans에 의해 양배추(Brassica oleracea var. capitata) 에 발병하는 심각한 병해이다. 최근, 양배추의 시들음병 저항 성 유전자, FocBo1, 과 연관된 분자 마커 (MTK-C)에 대한 보고가 있었다. 본 연구에서는, MTK-C를 이용하여, 국내 시 들음병 저항성 육종을 위한 양배추 육종 계통이 FocBo1 유 전자와 연관된 것인지 판별하고자 하였다. 상업적으로 판매되 고 있는 ‘대박나’를 대조군으로 하여 6가지의 양배추 육종계 통을 통제된 실험실 조건에서, 시들음병을 처리하고, 생물검 정에 따른 발병도와 식물체내 병원균의 생체량을 확인하여 육종계통의 시들음병에 대한 반응성을 정확히 평가 하였다. 그결과, KR-518, OK-517은 반복 실험을 통해 병징 뿐만 아 니라 병원균의 바이오 매스도 거의 검출되지 않는 강한 저항 성을 나타냈고, RK-P6-1, MT-624는 실험실내 병 처리 환경 에 따라 그 저항성과 감수성의 정도가 달라지는 각각 중도 저 항성, 감수성으로 분류 할 수 있으며, JK-2, HY-164는 병징 과 바이오매스 모두에서 병에 대한 감수성을 보임을 확인 하 였다. FocBo1 간의 상관관계를 알기 위하여, 6개 양배추 계 통의 genomic DNA를 추출하고, MTK-C 마커를 이용하여 PCR을 수행한 결과, 흥미롭게도 감수성 유전자원인 HY-164 식물체에서도 FocBo1 밴드가 관찰 되었다. 염기서열의 차이 를 확인하고자 증폭된 밴드를 클로닝하여 sequencing 한 결 과 감수성 HY-164와 저항성 KR-518 간의 차이는 없었다. 하지만, HY-164의 식물체에서는 FocBo1 유전자의 발현이 없는 것을 cDNA를 이용한 RT-PCR 결과로 확인 하였으며, 더하여, FocBo1 유전자의 발현 정도가 완전 저항성, 중도 저 항성을 결정하는 중요 요인이라는 것을 실험을 통해 확인 할 수 있었다. 더하여, 확인된 MTK-C 밴드의 염기서열을 이용 하여 단일밴드로 정확히 FocBo1 발현을 확인할 수 있는 프 라이머, FocBo1-C를 만들 수 있었다. 본 실험을 통하여, 국 내에서 보유한 6개 시들음병 관련 양배추 육종계통의 정확한 병 반응성을 정리 하였으며, 또한 이들의 시들음병 저항성이 FocBo1 유전자 및 발현과 밀접한 관련이 있다는 것을 확인 할 수 있었다. 따라서, FocBo1을 이용한 분자육종에서 저항 성과 감수성 판별을 위하여 genome 상의 FocBo1 존재뿐만 아니라 그 발현의 정도도 같이 확인 하여야 한다는 것을 확인 하였다.
사 사
본 연구는 농림축산식품부 양배추류 유전체 육종 통합 지 원 시스템 개발사업(관리번호: 610008-05)과 한국생명공학연 구원 기관 고유사업 지원에 의해 수행되었습니다. 본 연구 수 행에 있어 6가지의 양배추 육종 계통을 제공해주신 아시아 종 묘㈜와 Foc균주를 제공해주신 최경자 박사님께 감사드립니다.
References
- 1. Baik SY, Kim JC, Jang KS, Choi YH, Choi GJ. Development of efficient screening method for resistance of cabbage to Fusarium oxysporum f. sp. conglutinans. Res. Plant Dis 2011. 17: 13-18.
- 2. Blank LM. Fusarium resistance in Wisconsin all seasons cabbage. J. Agric. Res 1937. 55: 407-510.
- 3. Brown DW, Proctor RH. Fusarium : genomics, molecular and cellular biology 2013. Caister Academic Press. Norfolk.
- 4. Lee JH, Jo EJ, Jang KS, Choi YH, Kim JC, Choi GJ. Evaluation of cabbage- and broccoli-genetic resources for resistance to clubroot and Fusarium wilt. Res. Plant Dis 2014. 20: 235-244.
- 5. Li E, Wang G, Yang Y, Xiao J, Mao Z, Xie B. Microscopic analysis of the compatible and incompatible interactions between Fusarium oxysporum f. sp. conglutinans and cabbage. Eur. J. Plant Pathol 2015. 141: 597-609.
- 6. Michielse CB, Rep M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol 2009. 10: 311-324.
- 7. Moon YK, Kim WJ, Cho WD, Sung JM. Occurrence of Fusarium wilt on cruciferous vegetable crops and pathogenic differentiation of the causal fungus. Res. Plant Dis 2001. 7: 93-101.
- 8. Pu ZJ, Shimizu M, Zhang YJ, Nagaoka T, Hayashi T, Hori H, Matsumoto S, Fujimoto R, Okazaki K. Genetic mapping of a fusarium wilt resistance gene in Brassica oleracea. Mol. Breed 2012. 30: 809-818.
- 9. Shimizu M, Fujimoto R, Ying H, Pu ZJ, Ebe Y, Kawanabe T, Saeki N, Taylor JM, Kaji M, Dennis ES. Identification of candidate genes for fusarium yellows resistance in Chinese cabbage by differential expression analysis. Plant Mol. Biol 2014. 85: 247-257.
- 10. Shimizu M, Pu ZJ, Kawanabe T, Kitashiba H, Matsumoto S, Ebe Y, Sano M, Funaki T, Fukai E, Fujimoto R. Map-based cloning of a candidate gene conferring Fusarium yellows resistance in Brassica oleracea. Theor. Appl. Genet 2015. 128: 119-130.
- 11. Smith R. Effect of environmental factors upon, the resistance of cabbage ta yellows. J. Agric. Res 1930. 41: 1-15.
- 12. Song JH, Kim YW, Cho JY. Varietal difference and inheritance of cabbage yellows (Fusarium oxysporum f. sp. conglutinans Synder et Hansen) resistance in
cabbage. Korean J. Breed. Sci 1996. 28: 171-177.
- 13. Thatcher LF, Manners JM, Kazan K. Fusarium oxysporum hijacks COI1 mediated jasmonate signaling to promote disease development inArabidopsis. Plant J 2009. 58: 927-939.
- 14. TrusovY. ChakravortyD. BotellaJ. Fusarium oxysporum Infection assays in Arabidopsis. Running MP. G Protein-Coupled Receptor Signaling in Plants Clifton, NJ, Humana Press. 2013. 67-72.
- 15. Walker JC. Cauliflower, cabbage, and others. In Plant Diseases. The yearbook of agriculture 1953 1953. US Department of Agriculture. Washington, 425-430.
- 16. Walker JC, Wellman FL. Development of Three Midseason Varieties of Cabbage resistant to Yellows (Fusarium con-glutinans Woll. . J. Agric. Res 1927. 35: 785-809.