1 농촌진흥청 국립식량과학원
1 National Institute of Crop Science, RDA, Wanju 55365, Korea
2 농촌진흥청 국립식량과학원 중부작물부
2 Department of Central Area, NICS, RDA, Suweon 16429, Korea
3 충남대학교 농업생명대학
3 Department of Agronomy, Chungnam National University, Daejeon 34134, Korea
© Korean Society of Breeding Science All rights reserved
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
| Anther cultured (no.) | Cali induced (no. and %) | Plant regeneration (no. and %)z | Green plants (no. and %)y | Albino (no. and %)x | Doubled haploid lines (no.)w |
|---|---|---|---|---|---|
|
|
|||||
| 10,600 | 3,528(33.3) | 450(12.8) | 245(54.4) | 215(45.6) | 220 |
zPercent plant regeneration is the number of plants regenerated over number of cali plated
yPercent green is the number of green plants regenerated over total plants regenerated
xPercent albino is the number of albino plants regenerated over total plants regenerated
wDouble haploid lines are harvested plants having fertile grain
| Traitsz | PLy | TNN | NR | NS | NPr | NSPr | MNSPr | NSr | NSSr | MNSSr | Sr/Pr | SSr/SPr | CL | FL | PN | TGW | RRG | Yield |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (x1) | (x2) | (x3) | (x4) | (x5) | (x6) | (x7) | (x8) | (x9) | (x10) | (x11) | (x12) | (x13) | (x14) | (x15) | (x16) | (x17) | (x18) | |
|
|
||||||||||||||||||
| x1 | 0.547** | 0.472** | 0.515** | 0.544** | 0.589** | 0.354** | 0.415** | 0.396** | 0.229** | 0.241** | 0.209** | 0.209** | 0.556** | -0.392** | 0.381** | -0.210** | 0.198* | |
| x2 | 0.752** | 0.858** | 0.884** | 0.655** | 0.674** | 0.294** | 0.816** | 0.803** | 0.549** | 0.629** | 0.602** | 0.005ns | 0.513** | -0.325** | 0.159* | -0.313** | 0.376** | |
| x3 | 0.706** | 0.833** | 0.987** | 0.571** | 0.571** | 0.188* | 0.988** | 0.965** | 0.587** | 0.859** | 0.823** | -0.052ns | 0.412** | -0.302** | 0.044ns | -0.364** | 0.468** | |
| x4 | 0.659** | 0.846** | 0.990** | 0.609** | 0.630** | 0.286** | 0.967** | 0.958** | 0.640** | 0.820** | 0.793** | -0.039ns | 0.442** | -0.297** | 0.059ns | -0.378** | 0.456** | |
| x5 | 0.794** | 0.765** | 0.765** | 0.817** | 0.966** | 0.218** | 0.437** | 0.367** | 0.002ns | 0.075ns | 0.031ns | 0.194* | 0.476** | -0.343** | 0.395** | -0.156* | 0.118ns | |
| x6 | 0.803** | 0.745** | 0.746** | 0.812** | 0.979** | 0.462** | 0.443** | 0.380** | 0.050ns | 0.093ns | 0.033ns | 0.224** | 0.515** | -0.347** | 0.388** | -0.170* | 0.126ns | |
| x7 | 0.561** | 0.424** | 0.423** | 0.516** | 0.579** | 0.732** | 0.165* | 0.169* | 0.185* | 0.086ns | 0.009ns | 0.171* | 0.325** | -0.128ns | 0.118ns | -0.098ns | 0.07ns | |
| x8 | 0.594** | 0.799** | 0.991** | 0.969** | 0.674** | 0.656** | 0.367** | 0.988** | 0.643** | 0.926** | 0.896** | -0.094ns | 0.361** | -0.266** | -0.026ns | -0.370** | 0.491** | |
| x9 | 0.580** | 0.792** | 0.982** | 0.966** | 0.649** | 0.633** | 0.360** | 0.994** | 0.744** | 0.942** | 0.933** | -0.130ns | 0.335** | -0.226** | -0.073ns | -0.388** | 0.497** | |
| x10 | 0.401** | 0.610** | 0.739** | 0.759** | 0.418** | 0.417** | 0.281** | 0.762** | 0.821** | 0.713** | 0.785** | -0.267** | 0.180* | 0.010ns | -0.283** | -0.347** | 0.382** | |
| x11 | 0.298** | 0.590** | 0.825** | 0.770** | 0.288** | 0.256** | 0.058ns | 0.888** | 0.907** | 0.806** | 0.986** | -0.182* | 0.204** | -0.164ns | -0.194* | -0.354** | 0.495** | |
| x12 | 0.346** | 0.620** | 0.862** | 0.807** | 0.338** | 0.324** | 0.161* | 0.921** | 0.927** | 0.757** | 0.986** | -0.222** | 0.166* | -0.119ns | -0.234** | -0.369** | 0.492** | |
| x13 | 0.061ns | -0.160* | -0.221** | -0.194ns | 0.060ns | 0.079ns | 0.114ns | -0.266** | -0.293** | -0.424** | -0.415** | -0.376** | -0.075ns | -0.047ns | 0.386** | 0.045ns | -0.100ns | |
| x14 | 0.549** | 0.432** | 0.413** | 0.421** | 0.420** | 0.426** | 0.289** | 0.388** | 0.369** | 0.189* | 0.237** | 0.279** | -0.006ns | -0.418** | 0.285** | -0.163* | 0.241** | |
| x15 | -0.807** | -0.762** | -0.733** | -0.750** | -0.726** | -0.713** | -0.417** | -0.693** | -0.678** | -0.498** | -0.478** | -0.515** | 0.148ns | -0.548** | -0.361** | 0.091ns | 0.142ns | |
| x16 | 0.314** | 0.043ns | -0.088ns | -0.055ns | 0.200* | 0.194* | 0.107ns | -0.142ns | -0.158* | -0.252** | -0.310** | -0.291** | 0.322** | 0.375** | -0.180* | 0.405** | -0.080ns | |
| x17 | -0.329** | -0.398** | -0.538** | -0.527** | -0.321** | -0.308** | -0.162* | -0.552** | -0.562** | -0.511** | -0.550** | -0.545** | 0.317** | -0.164* | 0.378** | 0.458** | -0.169* | |
| x18 | 0.196* | 0.261** | 0.410** | 0.413** | 0.270** | 0.271** | 0.187* | 0.415** | 0.427** | 0.427** | 0.402** | 0.400** | -0.14ns | 0.254** | -0.202** | 0.051ns | -0.323** | |
zUpper and lower diagonal indicate the value in 2011 and 2012, respectively
yAbbreviations are listed in table 1
| Trait | 2011 | 2012 | ||||||
|---|---|---|---|---|---|---|---|---|
|
|
||||||||
| Eigenvector | Correlation coefficient | Eigenvector | Correlation coefficient | |||||
|
|
||||||||
| Prin1z | Prin2 | Prin1 | Prin2 | Prin1z | Prin2 | Prin1 | Prin2 | |
|
|
||||||||
| PLy | 0.197 | 0.277 | 0.567** | 0.511** | 0.236 | 0.299 | 0.743** | 0.516** |
| TNN | 0.311 | 0.102 | 0.895** | 0.189* | 0.276 | 0.092 | 0.870** | 0.159* |
| NR | 0.339 | -0.015 | 0.975** | -0.028ns | 0.310 | -0.036 | 0.979** | -0.062ns |
| NS | 0.343 | 0.012 | 0.987** | 0.023ns | 0.313 | 0.010 | 0.987** | 0.017ns |
| NPr | 0.193 | 0.369 | 0.554** | 0.680** | 0.249 | 0.280 | 0.786** | 0.484** |
| NSPr | 0.202 | 0.385 | 0.582** | 0.709** | 0.248 | 0.303 | 0.781** | 0.523** |
| MNSPr | 0.102 | 0.191 | 0.292** | 0.352** | 0.158 | 0.276 | 0.498** | 0.476** |
| NSr | 0.335 | -0.086 | 0.964** | -0.158* | 0.305 | -0.099 | 0.963** | -0.171* |
| NSSr | 0.334 | -0.128 | 0.960** | -0.235** | 0.305 | -0.121 | 0.962** | -0.209** |
| MNSSr | 0.235 | -0.252 | 0.676** | -0.465** | 0.247 | -0.212 | 0.779** | -0.366** |
| Sr/Pr | 0.292 | -0.248 | 0.841** | -0.456** | 0.249 | -0.316 | 0.787** | -0.545** |
| SSr/SPr | 0.284 | -0.285 | 0.816** | -0.525** | 0.260 | -0.275 | 0.820** | -0.474** |
| CL | -0.023 | 0.256 | -0.067ns | 0.472** | -0.079 | 0.330 | -0.249** | 0.570** |
| FL | 0.178 | 0.236 | 0.511** | 0.436** | 0.153 | 0.220 | 0.484** | 0.381** |
| PN | -0.116 | -0.235 | -0.333** | -0.434** | -0.254 | -0.167 | -0.801** | -0.288** |
| TGW | 0.016 | 0.394 | 0.045ns | 0.726** | -0.017 | 0.416 | -0.053ns | 0.718** |
| RRG | -0.148 | 0.110 | -0.425** | 0.203** | -0.184 | 0.215 | -0.580** | 0.370** |
| Yield | 0.176 | -0.133 | 0.507** | -0.245** | 0.141 | -0.072 | 0.446** | -0.125ns |
| Eigen value | 8.27 | 3.40 | 9.95 | 2.98 | ||||
| Contribution (%) | 46.0 | 18.9 | 55.3 | 16.6 | ||||
| Cumulative contribution (%) | 46.0 | 64.8 | 55.3 | 71.8 | ||||
| Group | n | PLz (cm) | TNN (mm) | NR | NS | NPr | NSPr | MNSPr | NSr | NSSr | MNSSr | Sr/Pr | SSr/SPr | CL (cm) | FL (cm) | PN | TGW (g) | RRG (%) | Yield (g) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||
| Deuraechan | 3 | 26.3ay | 1.74a | 42a | 166a | 14a | 86a | 6.1a | 28a | 80a | 2.9a | 1.9ab | 0.9bc | 69.1a | 32.4a | 9c | 25.0a | 87.3ab | 535bc |
| Boramchan | 3 | 21.0d | 1.53bc | 35bc | 129b | 11d | 60d | 5.7d | 24ab | 69ab | 2.8ab | 2.3a | 1.1a | 67.6a | 30.3b | 11a | 24.3a | 91.3a | 551b |
| a | 108 | 24.7ab | 1.63ab | 40ab | 156a | 13b | 79b | 6.0ab | 27a | 78a | 2.9a | 2.1ab | 1.0abc | 68.7a | 32.3a | 9bc | 24.2a | 84.1b | 520c |
| (20.1~27.1) | (1.29~1.88) | (26~51) | (107~197) | (11~15) | (65~93) | (5.7~6.3) | (15~38) | (41~118) | (2.6~3.1) | (1.3~3.1) | (0.6~1.6) | (62.3~74.3) | (28.3~35.7) | (8~13) | (21.7~27.2) | (74.1~93.0) | (477~554) | ||
| b | 42 | 22.6cd | 1.49c | 33c | 127b | 12c | 69c | 5.8c | 21b | 58b | 2.7b | 1.8b | 0.8c | 69.5a | 31.0ab | 11ab | 24.0a | 87.3ab | 467d |
| (19.2~26.5) | (1.32~1.77) | (24~44) | (97~172) | (10~14) | (57~85) | (5.6~6.1) | (13~31) | (35~90) | (2.5~3) | (1.2~2.6) | (0.5~1.3) | (63.5~75.0) | (26.9~33.5) | (9~12) | (21.0~27.1) | (69.4~94.9) | (410~501) | ||
| c | 13 | 23.7bc | 1.59bc | 40ab | 156a | 13bc | 75bc | 5.9bc | 28a | 81a | 2.9a | 2.2a | 1.1ab | 68.2a | 31.8ab | 10abc | 24.2a | 83.9c | 581a |
| (20.7~26.5) | (1.25~1.80) | (27~46) | (108~179) | (11~14) | (62~85) | (5.7~6.1) | (16~33) | (42~102) | (2.7~3.1) | (1.4~2.7) | (0.6~1.4) | (63.9~72.1) | (28.8~35.0) | (8~13) | (21.9~25.7) | (75.2~92.4) | (555~629) | ||
| Total | 163 | 24.1 | 1.59 | 38.2 | 149 | 13 | 76 | 5.9 | 26 | 73 | 2.8 | 2.0 | 1.0 | 68.8 | 31.9 | 10 | 24.1 | 84.9 | 512 |
| C.V.(%)x | 163 | 8.3 | 8.2 | 15.4 | 15.1 | 8.7 | 10.3 | 3.4 | 20.0 | 23.0 | 3.6 | 15.0 | 20.0 | 3.5 | 5.0 | 11.3 | 4.5 | 5.3 | 7.2 |
zAbbreviations are listed in table 1
yMeans with same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
xC.V. = coefficient of variation
| Var./lines | PLz (cm) | TNN | NR | NS | NPr | NSPr | MNSPr | NSr | NSSr | MNSSr | Sr/Pr | SSr/SPr | CL (cm) | FL | PN | TGW (g) | RRG (%) | MRY (kg/10a) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||||||||
| Deuraechan (DR) | 26aby | 1.76ab | 32cde | 122bcd | 11abcd | 67bcd | 5.9bcde | 20de | 55efgh | 2.7ef | 1.7ef | 0.8ghi | 73ab | 36.0ab | 10efg | 25.3bc | 91.2a | 512b |
| Boramchan (BR) | 21e | 1.50def | 31cde | 111de | 9h | 51h | 5.5h | 22cde | 60def | 2.8cde | 2.3abc | 1.2bc | 71bcd | 32.1de | 13abc | 24.0ef | 91.4a | 567ab |
| HR28832-AC60-1-1 | 23cd | 1.62bcde | 31cde | 117bcde | 10fgh | 56gh | 5.8defg | 21cde | 61def | 2.8c | 2.2bcd | 1.1cd | 72bc | 34.2abcde | 12bcd | 23.7fg | 91.9a | 596a |
| (AC60) | ||||||||||||||||||
| HR28832-AC105-1-1 | 24cd | 1.69abc | 35abc | 132abc | 11bcde | 65cde | 5.8cdefg | 23bcd | 67bcde | 2.8c | 2.0de | 1.0def | 68e | 33.5bcde | 12abcd | 24.3e | 84.1bcd | 559ab |
| (AC105) | ||||||||||||||||||
| HR28832-AC107-1-1 | 26ab | 1.68abc | 39a | 149a | 12a | 73a | 6.0ab | 27ab | 75ab | 2.8cd | 2.2bcd | 1.0de | 73ab | 37.2a | 9h | 25.7ab | 83.8bcd | 516b |
| (AC107) | ||||||||||||||||||
| HR28832-AC109-1-1 | 23de | 1.34g | 26f | 101e | 10gh | 57g | 5.9abc | 16f | 44h | 2.7ef | 1.6fg | 0.7hi | 72bc | 31.0e | 13ab | 24.5de | 89.4ab | 561ab |
| (AC109) | ||||||||||||||||||
| HR28832-AC112-1-1 | 23de | 1.47efg | 31cde | 117bcde | 11def | 60gef | 5.7fg | 21de | 57efg | 2.7def | 1.9def | 0.9efg | 71bcd | 32.0de | 12cde | 23.2g | 92.2a | 549ab |
| (AC112) | ||||||||||||||||||
| HR28832-AC123-1-1 | 27a | 1.62bcde | 34abc | 133ab | 12abc | 71abc | 6.0a | 23cd | 62cdef | 2.8cdef | 1.9def | 0.9fgh | 73abc | 32.6cde | 10gh | 25.7ab | 85.3bcd | 541ab |
| (AC123) | ||||||||||||||||||
| HR28832-AC130-1-1 | 25ab | 1.66abc | 29ef | 115cde | 12abcd | 67bcd | 5.8cdef | 18ef | 47gh | 2.7ef | 1.5g | 0.7i | 73ab | 34.1abcde | 10fgh | 25.5abc | 87.6abc | 516b |
| (AC130) | ||||||||||||||||||
| HR28832-AC134-1-1 | 26ab | 1.58cdef | 37ab | 143a | 12ab | 72ab | 6.0a | 25abc | 71abcd | 2.8bc | 2.0cd | 1.0def | 75a | 35.8abc | 10gh | 25.0cd | 82.7cd | 500b |
| (AC134) | ||||||||||||||||||
| HR28832-AC149-1-1 | 23cd | 1.62bcde | 35abc | 132abc | 10efg | 58gf | 5.7g | 25abc | 74abc | 2.9ab | 2.4ab | 1.2ab | 68de | 31.3e | 11cdef | 23.3g | 85.0bcd | 564ab |
| (AC149) | ||||||||||||||||||
| HR28832-AC152-1-1 | 23cd | 1.65abcd | 38a | 148a | 11cde | 66cde | 6.0ab | 27ab | 82a | 3.0a | 2.4ab | 1.2ab | 72bcd | 35.9ab | 11cdef | 24.0ef | 89.5ab | 595a |
| (AC152) | ||||||||||||||||||
| HR28832-AC156-1-1 | 22de | 1.46fg | 29def | 110de | 10fgh | 56g | 5.7efg | 20def | 54fgh | 2.7cdef | 2.0de | 0.9defg | 71bcd | 35.5abc | 14a | 23.7fg | 90.9a | 590a |
| (AC156) | ||||||||||||||||||
| HR28832-AC161-1-1 | 25bc | 1.73abc | 38a | 146a | 11de | 63def | 5.9bcd | 28a | 83a | 3.0a | 2.6a | 1.3a | 69de | 35.1abcd | 11defg | 25.5abc | 84.9bcd | 605a |
| (AC161) | ||||||||||||||||||
| HR28832-AC178-1-1 | 25abc | 1.80a | 33bcde | 122bcd | 11cde | 63def | 5.8cdefg | 22cd | 58efg | 2.7f | 2.0de | 0.9defg | 70cde | 33.8bcde | 10gh | 26.1a | 80.7d | 507b |
| (AC178) | ||||||||||||||||||
| Mean | 24 | 1.61 | 33 | 125 | 11 | 62 | 5.8 | 62 | 22 | 2.8 | 2.0 | 1.0 | 71 | 34 | 11 | 24.6 | 87.5 | 552 |
| C.V. | 7.5 | 8.7 | 27.3 | 28.0 | 18.2 | 21.0 | 5.2 | 36.4 | 40.3 | 7.1 | 30.0 | 30.0 | 3.5 | 6.9 | 13.5 | 4.1 | 5.3 | 8.2 |
zAbbreviations are listed in table 1
yMeans with same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
| Traits | PLz | TNN | NR | NS | NPr | NSPr | MNSPr | NSr | NSSr | MNSSr | Sr/Pr | SSr/SPr | CL | FL | PN | TGW | RRG | MRY |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (x1) | (x2) | (x3) | (x4) | (x5) | (x6) | (x7) | (x8) | (x9) | (x10) | (x11) | (x12) | (x13) | (x14) | (x15) | (x16) | (x17) | (x18) | |
|
|
||||||||||||||||||
| x1 | 0.612* | 0.464ns | 0.558* | 0.902** | 0.907** | 0.728** | 0.279ns | 0.218ns | -0.008ns | -0.265ns | -0.213ns | 0.420ns | 0.473ns | -0.918** | 0.802** | -0.531* | -0.647** | |
| x2 | 0.568* | 0.564* | 0.584* | 0.519* | 0.209ns | 0.483ns | 0.443ns | 0.251ns | 0.197ns | 0.210ns | -0.233ns | 0.486ns | -0.684** | 0.571* | -0.457ns | -0.289ns | ||
| x3 | 0.984** | 0.581* | 0.582* | 0.423ns | 0.975** | 0.952** | 0.761** | 0.647** | 0.692** | -0.109ns | 0.523* | -0.570* | 0.232ns | -0.518* | 0.010ns | |||
| x4 | 0.671** | 0.687** | 0.560* | 0.932** | 0.914** | 0.739** | 0.543* | 0.588* | -0.023ns | 0.554* | -0.619* | 0.301ns | -0.532* | -0.040ns | ||||
| x5 | 0.987** | 0.713** | 0.384ns | 0.320ns | 0.059ns | -0.232ns | -0.182ns | 0.341ns | 0.549* | -0.882** | 0.660** | -0.548* | -0.650** | |||||
| x6 | 0.815** | 0.390ns | 0.334ns | 0.092ns | -0.232ns | -0.175ns | 0.393ns | 0.558* | -0.847** | 0.663** | -0.539* | -0.591* | ||||||
| x7 | 0.285ns | 0.272ns | 0.164ns | -0.212ns | -0.143ns | 0.450ns | 0.463ns | -0.536* | 0.541* | -0.375ns | -0.247ns | |||||||
| x8 | 0.992** | 0.847** | 0.798** | 0.835** | -0.217ns | 0.442ns | -0.404ns | 0.082ns | -0.437ns | 0.190ns | ||||||||
| x9 | 0.907** | 0.835** | 0.861** | -0.249ns | 0.407ns | -0.330ns | 0.021ns | -0.390ns | 0.278ns | |||||||||
| x10 | 0.871** | 0.857** | -0.301ns | 0.244ns | -0.047ns | -0.211ns | -0.164ns | 0.540* | ||||||||||
| x11 | 0.992** | -0.471ns | 0.102ns | 0.108ns | -0.318ns | -0.103ns | 0.598* | |||||||||||
| x12 | -0.426ns | 0.157ns | 0.060ns | -0.269ns | -0.148ns | 0.563* | ||||||||||||
| x13 | 0.354ns | -0.328ns | 0.249ns | 0.187ns | -0.468ns | |||||||||||||
| x14 | -0.466ns | 0.380ns | -0.160ns | -0.158ns | ||||||||||||||
| x15 | -0.743** | 0.551* | 0.679** | |||||||||||||||
| x16 | -0.579* | -0.584* | ||||||||||||||||
| x17 | 0.409ns | |||||||||||||||||
zAbbreviations are listed in table 1
| Trait | Eigenvector | Correlation coefficient | ||||
|---|---|---|---|---|---|---|
|
|
||||||
| Prin1z | Prin2 | Prin3 | Prin1 | Prin2 | Prin3 | |
|
|
||||||
| PL | 0.275 | -0.230 | -0.011 | 0.789** | -0.551* | -0.014ns |
| TNN | 0.246 | -0.015 | -0.375 | 0.707** | -0.037ns | -0.451ns |
| NR | 0.313 | 0.172 | 0.030 | 0.898** | 0.412ns | 0.036ns |
| NS | 0.326 | 0.127 | 0.087 | 0.937** | 0.306ns | 0.105ns |
| NPr | 0.292 | -0.198 | 0.027 | 0.838** | -0.476ns | 0.032ns |
| NSPr | 0.293 | -0.195 | 0.108 | 0.842** | -0.468ns | 0.130ns |
| MNSPr | 0.228 | -0.150 | 0.342 | 0.653** | -0.359ns | 0.411ns |
| NSr | 0.275 | 0.249 | 0.027 | 0.788** | 0.598* | 0.032ns |
| NSSr | 0.260 | 0.274 | 0.053 | 0.745** | 0.659** | 0.064ns |
| MNSSr | 0.176 | 0.329 | 0.145 | 0.506ns | 0.791** | 0.174ns |
| Sr/Pr | 0.103 | 0.389 | -0.036 | 0.297ns | 0.933** | -0.043ns |
| SSr/SPr | 0.123 | 0.377 | -0.014 | 0.352ns | 0.906** | -0.016ns |
| CL | 0.040 | -0.245 | 0.569 | 0.114ns | -0.589* | 0.684** |
| FL | 0.221 | -0.034 | 0.313 | 0.635* | -0.082ns | 0.376ns |
| PN | -0.287 | 0.178 | 0.112 | -0.825** | 0.427ns | 0.134ns |
| TGW | 0.203 | -0.241 | -0.227 | 0.582* | -0.579* | -0.273ns |
| RRG | -0.225 | 0.044 | 0.422 | -0.645** | 0.105ns | 0.507ns |
| MRY | -0.109 | 0.331 | 0.181 | -0.312ns | 0.795** | 0.218ns |
| Eigen value | 8.24 | 5.77 | 1.45 | |||
| Contribution (%) | 45.8 | 32.0 | 0.08 | |||
| Cumulative contribution (%) | 45.8 | 77.8 | 85.8 | |||
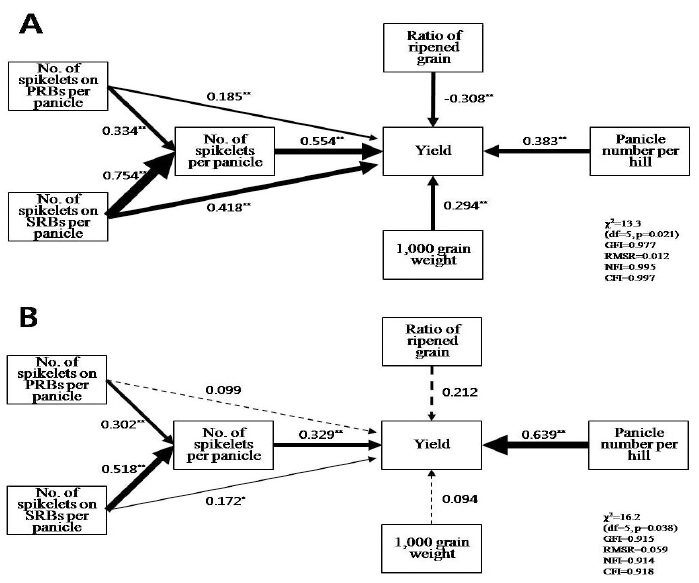
| Var./lines | Heading date (DASz) | CL (cm) | PL (cm) | PN | NS | RRG (%) | TGW (g) | RRY (kg/10a) | BRR (%) | BRY (kg/10a) | MRY (kg/10a) | Index of MRY (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||
| Deuraechan (DR) | 107ay | 69a | 24a | 9c | 142a | 91.1ab | 25.8a | 641b | 81.6b | 523b | 481b | 100 |
| Boramchan (BR) | 108a | 69a | 20c | 12a | 108b | 94.5a | 24.4b | 736a | 82.5ab | 607a | 558a | 116 |
| HR28832-AC60-1-1 | 108a | 66bc | 22b | 11b | 129ab | 91.0ab | 24.5b | 772a | 81.0b | 625a | 575a | 119 |
| (AC60) | ||||||||||||
| HR28832-AC152-1-1 | 108a | 68ab | 22b | 11b | 140a | 90.5ab | 24.7b | 744a | 81.6b | 607a | 558a | 116 |
| (AC152) | ||||||||||||
| HR28832-AC156-1-1 | 108a | 64c | 20c | 12a | 108b | 94.8a | 24.0c | 741a | 83.2a | 617a | 567a | 118 |
| (AC156) | ||||||||||||
| HR28832-AC161-1-1 | 106a | 65bc | 22b | 11b | 143a | 86.3b | 25.9a | 748a | 81.1b | 607a | 559a | 116 |
| (AC161) | ||||||||||||
zAbbreviations are listed in table 1
yMeans with same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
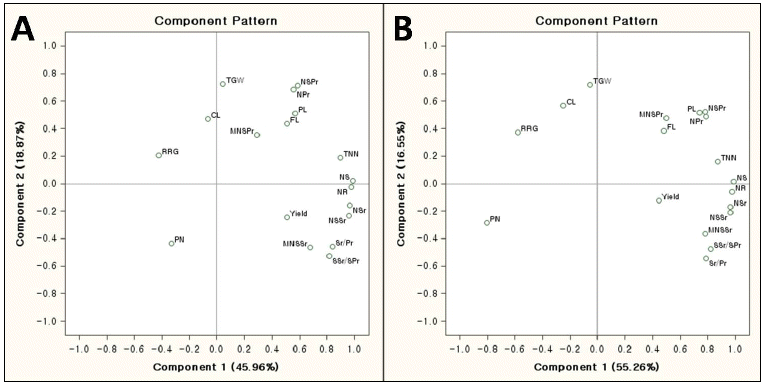
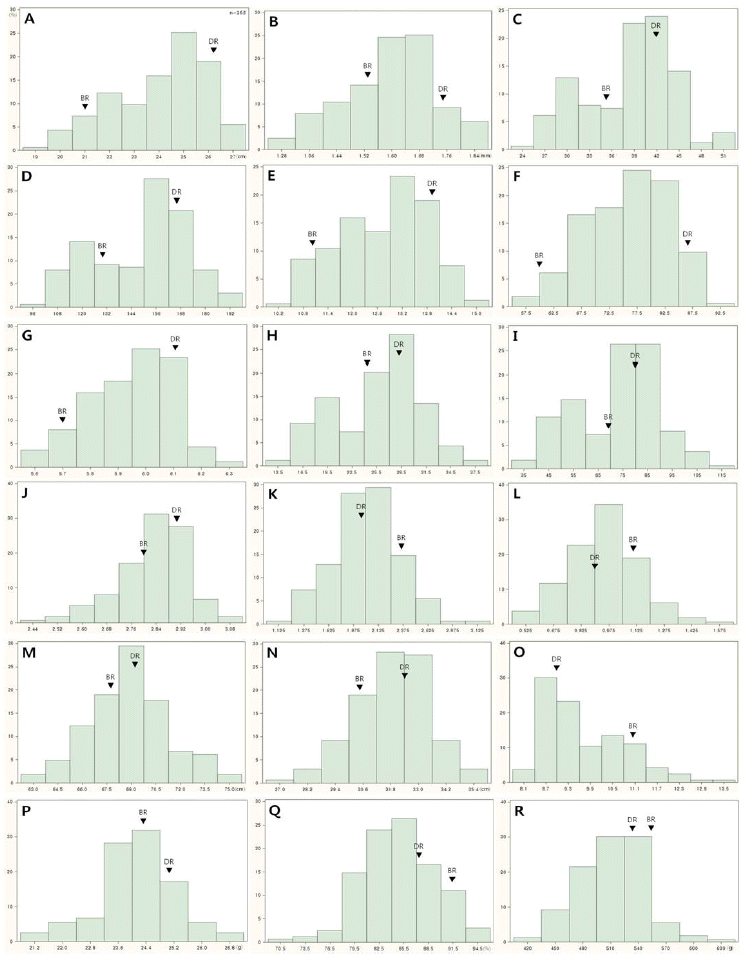

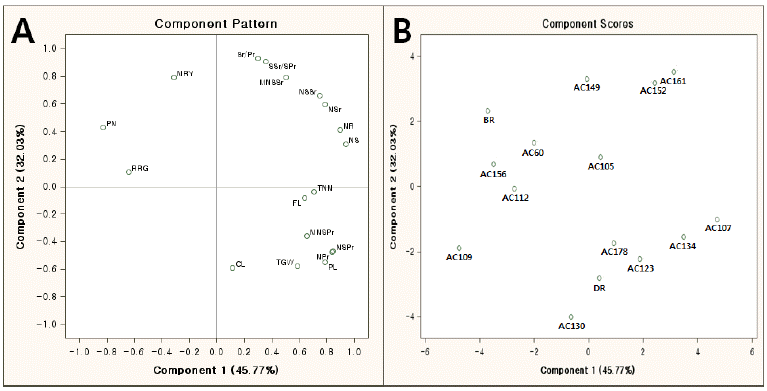
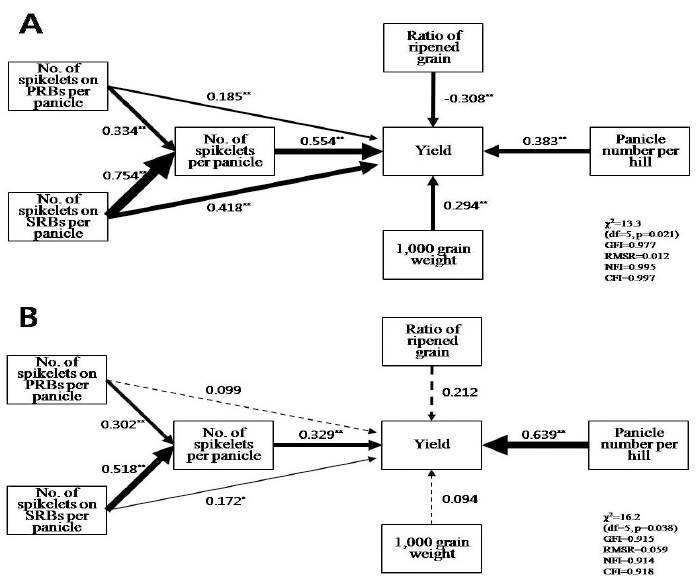
The abbreviations for panicle and yield-related traits.
| Abbreviations: | |
|---|---|
|
|
|
| PL | Panicle length |
| TNN | Thickness of neck node |
| PRB | Primary rachis-branch |
| SRB | Secondary rachis-branch |
| NR | Number of rachis-branches per panicle |
| NS | Number of spikelets per panicle |
| NPr | Number of PRBs per panicle |
| NSPr | Number of spikelets on PRBs per panicle |
| MNSPr | Mean number of spikelets on a PRB |
| NSr | Number of SRBs per panicle |
| NSSr | Number of spikelets on SRBs per panicle |
| MNSSr | Mean number of spikelets on a SRB |
| Sr/Pr | Ratio of SRBs to PRBs |
| SSr/SPr | Ratio of spikelets on SRBs to spikelets on PRBs |
| CL | Culm length |
| FL | Flag leaf length |
| PN | Panicle number per hill |
| TGW | 1,000 grain weight |
| RRG | Ratio of ripened grain |
| BRR | Brown/rough rice ratio |
| RRY | Rough rice yield |
| BRY | Brown rice yield |
| MRY | Milled rice yield |
| DAS | Day after seedling |
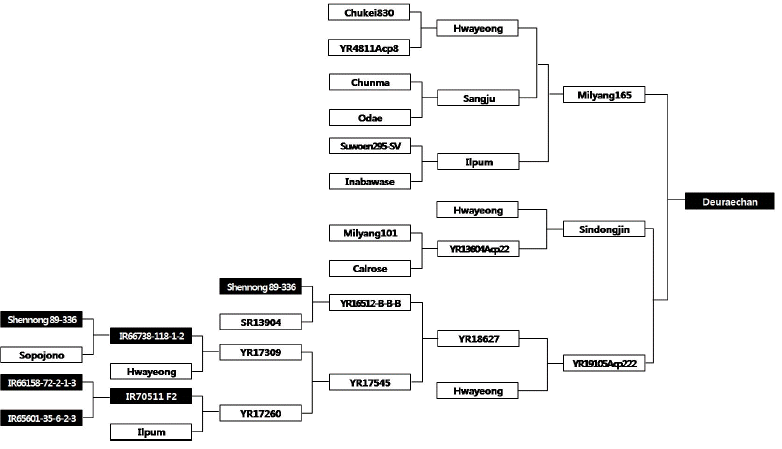
Anther culture ability and doubled haploid lines derived from the cross of Deuraechan/Boramchan.
| Anther cultured (no.) | Cali induced (no. and %) | Plant regeneration (no. and %) |
Green plants (no. and %) |
Albino (no. and %) |
Doubled haploid lines (no.) |
|---|---|---|---|---|---|
|
|
|||||
| 10,600 | 3,528(33.3) | 450(12.8) | 245(54.4) | 215(45.6) | 220 |
zPercent plant regeneration is the number of plants regenerated over number of cali plated
yPercent green is the number of green plants regenerated over total plants regenerated
xPercent albino is the number of albino plants regenerated over total plants regenerated
wDouble haploid lines are harvested plants having fertile grain
Correlation coefficient among panicle and yield related traits in 2011 and 2012 (n=163).
| Traits |
PL |
TNN | NR | NS | NPr | NSPr | MNSPr | NSr | NSSr | MNSSr | Sr/Pr | SSr/SPr | CL | FL | PN | TGW | RRG | Yield |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (x1) | (x2) | (x3) | (x4) | (x5) | (x6) | (x7) | (x8) | (x9) | (x10) | (x11) | (x12) | (x13) | (x14) | (x15) | (x16) | (x17) | (x18) | |
|
|
||||||||||||||||||
| x1 | 0.547** | 0.472** | 0.515** | 0.544** | 0.589** | 0.354** | 0.415** | 0.396** | 0.229** | 0.241** | 0.209** | 0.209** | 0.556** | -0.392** | 0.381** | -0.210** | 0.198* | |
| x2 | 0.752** | 0.858** | 0.884** | 0.655** | 0.674** | 0.294** | 0.816** | 0.803** | 0.549** | 0.629** | 0.602** | 0.005ns | 0.513** | -0.325** | 0.159* | -0.313** | 0.376** | |
| x3 | 0.706** | 0.833** | 0.987** | 0.571** | 0.571** | 0.188* | 0.988** | 0.965** | 0.587** | 0.859** | 0.823** | -0.052ns | 0.412** | -0.302** | 0.044ns | -0.364** | 0.468** | |
| x4 | 0.659** | 0.846** | 0.990** | 0.609** | 0.630** | 0.286** | 0.967** | 0.958** | 0.640** | 0.820** | 0.793** | -0.039ns | 0.442** | -0.297** | 0.059ns | -0.378** | 0.456** | |
| x5 | 0.794** | 0.765** | 0.765** | 0.817** | 0.966** | 0.218** | 0.437** | 0.367** | 0.002ns | 0.075ns | 0.031ns | 0.194* | 0.476** | -0.343** | 0.395** | -0.156* | 0.118ns | |
| x6 | 0.803** | 0.745** | 0.746** | 0.812** | 0.979** | 0.462** | 0.443** | 0.380** | 0.050ns | 0.093ns | 0.033ns | 0.224** | 0.515** | -0.347** | 0.388** | -0.170* | 0.126ns | |
| x7 | 0.561** | 0.424** | 0.423** | 0.516** | 0.579** | 0.732** | 0.165* | 0.169* | 0.185* | 0.086ns | 0.009ns | 0.171* | 0.325** | -0.128ns | 0.118ns | -0.098ns | 0.07ns | |
| x8 | 0.594** | 0.799** | 0.991** | 0.969** | 0.674** | 0.656** | 0.367** | 0.988** | 0.643** | 0.926** | 0.896** | -0.094ns | 0.361** | -0.266** | -0.026ns | -0.370** | 0.491** | |
| x9 | 0.580** | 0.792** | 0.982** | 0.966** | 0.649** | 0.633** | 0.360** | 0.994** | 0.744** | 0.942** | 0.933** | -0.130ns | 0.335** | -0.226** | -0.073ns | -0.388** | 0.497** | |
| x10 | 0.401** | 0.610** | 0.739** | 0.759** | 0.418** | 0.417** | 0.281** | 0.762** | 0.821** | 0.713** | 0.785** | -0.267** | 0.180* | 0.010ns | -0.283** | -0.347** | 0.382** | |
| x11 | 0.298** | 0.590** | 0.825** | 0.770** | 0.288** | 0.256** | 0.058ns | 0.888** | 0.907** | 0.806** | 0.986** | -0.182* | 0.204** | -0.164ns | -0.194* | -0.354** | 0.495** | |
| x12 | 0.346** | 0.620** | 0.862** | 0.807** | 0.338** | 0.324** | 0.161* | 0.921** | 0.927** | 0.757** | 0.986** | -0.222** | 0.166* | -0.119ns | -0.234** | -0.369** | 0.492** | |
| x13 | 0.061ns | -0.160* | -0.221** | -0.194ns | 0.060ns | 0.079ns | 0.114ns | -0.266** | -0.293** | -0.424** | -0.415** | -0.376** | -0.075ns | -0.047ns | 0.386** | 0.045ns | -0.100ns | |
| x14 | 0.549** | 0.432** | 0.413** | 0.421** | 0.420** | 0.426** | 0.289** | 0.388** | 0.369** | 0.189* | 0.237** | 0.279** | -0.006ns | -0.418** | 0.285** | -0.163* | 0.241** | |
| x15 | -0.807** | -0.762** | -0.733** | -0.750** | -0.726** | -0.713** | -0.417** | -0.693** | -0.678** | -0.498** | -0.478** | -0.515** | 0.148ns | -0.548** | -0.361** | 0.091ns | 0.142ns | |
| x16 | 0.314** | 0.043ns | -0.088ns | -0.055ns | 0.200* | 0.194* | 0.107ns | -0.142ns | -0.158* | -0.252** | -0.310** | -0.291** | 0.322** | 0.375** | -0.180* | 0.405** | -0.080ns | |
| x17 | -0.329** | -0.398** | -0.538** | -0.527** | -0.321** | -0.308** | -0.162* | -0.552** | -0.562** | -0.511** | -0.550** | -0.545** | 0.317** | -0.164* | 0.378** | 0.458** | -0.169* | |
| x18 | 0.196* | 0.261** | 0.410** | 0.413** | 0.270** | 0.271** | 0.187* | 0.415** | 0.427** | 0.427** | 0.402** | 0.400** | -0.14ns | 0.254** | -0.202** | 0.051ns | -0.323** | |
zUpper and lower diagonal indicate the value in 2011 and 2012, respectively
yAbbreviations are listed in table
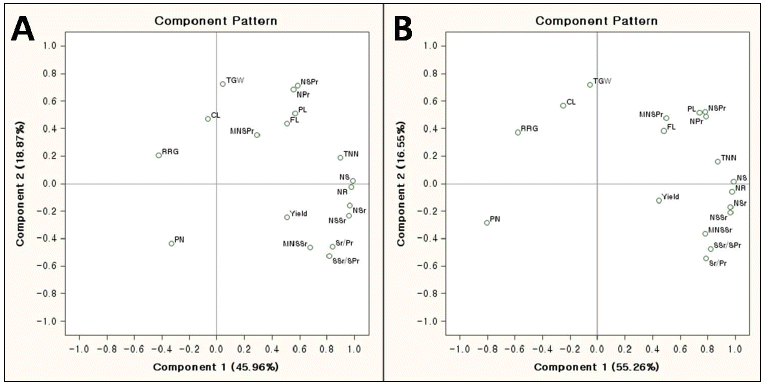
Eigenvectors for the principal components of traits and correlation coefficient between the traits and principal components.
| Trait | 2011 | 2012 | ||||||
|---|---|---|---|---|---|---|---|---|
|
|
||||||||
| Eigenvector | Correlation coefficient | Eigenvector | Correlation coefficient | |||||
|
|
||||||||
| Prin1 |
Prin2 | Prin1 | Prin2 | Prin1 |
Prin2 | Prin1 | Prin2 | |
|
|
||||||||
| PL |
0.197 | 0.277 | 0.567** | 0.511** | 0.236 | 0.299 | 0.743** | 0.516** |
| TNN | 0.311 | 0.102 | 0.895** | 0.189* | 0.276 | 0.092 | 0.870** | 0.159* |
| NR | 0.339 | -0.015 | 0.975** | -0.028ns | 0.310 | -0.036 | 0.979** | -0.062ns |
| NS | 0.343 | 0.012 | 0.987** | 0.023ns | 0.313 | 0.010 | 0.987** | 0.017ns |
| NPr | 0.193 | 0.369 | 0.554** | 0.680** | 0.249 | 0.280 | 0.786** | 0.484** |
| NSPr | 0.202 | 0.385 | 0.582** | 0.709** | 0.248 | 0.303 | 0.781** | 0.523** |
| MNSPr | 0.102 | 0.191 | 0.292** | 0.352** | 0.158 | 0.276 | 0.498** | 0.476** |
| NSr | 0.335 | -0.086 | 0.964** | -0.158* | 0.305 | -0.099 | 0.963** | -0.171* |
| NSSr | 0.334 | -0.128 | 0.960** | -0.235** | 0.305 | -0.121 | 0.962** | -0.209** |
| MNSSr | 0.235 | -0.252 | 0.676** | -0.465** | 0.247 | -0.212 | 0.779** | -0.366** |
| Sr/Pr | 0.292 | -0.248 | 0.841** | -0.456** | 0.249 | -0.316 | 0.787** | -0.545** |
| SSr/SPr | 0.284 | -0.285 | 0.816** | -0.525** | 0.260 | -0.275 | 0.820** | -0.474** |
| CL | -0.023 | 0.256 | -0.067ns | 0.472** | -0.079 | 0.330 | -0.249** | 0.570** |
| FL | 0.178 | 0.236 | 0.511** | 0.436** | 0.153 | 0.220 | 0.484** | 0.381** |
| PN | -0.116 | -0.235 | -0.333** | -0.434** | -0.254 | -0.167 | -0.801** | -0.288** |
| TGW | 0.016 | 0.394 | 0.045ns | 0.726** | -0.017 | 0.416 | -0.053ns | 0.718** |
| RRG | -0.148 | 0.110 | -0.425** | 0.203** | -0.184 | 0.215 | -0.580** | 0.370** |
| Yield | 0.176 | -0.133 | 0.507** | -0.245** | 0.141 | -0.072 | 0.446** | -0.125ns |
| Eigen value | 8.27 | 3.40 | 9.95 | 2.98 | ||||
| Contribution (%) | 46.0 | 18.9 | 55.3 | 16.6 | ||||
| Cumulative contribution (%) | 46.0 | 64.8 | 55.3 | 71.8 | ||||
zPrin1 and Prin2 indicate the first and second component, respectively
yAbbreviations are listed in table 1
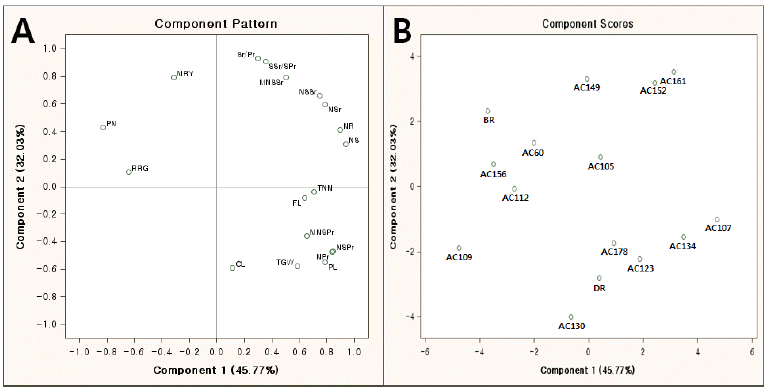
Average and range of traits value of the parents and groups divided by cluster analysis in 2012.
| Group | n | PL |
TNN (mm) | NR | NS | NPr | NSPr | MNSPr | NSr | NSSr | MNSSr | Sr/Pr | SSr/SPr | CL (cm) | FL (cm) | PN | TGW (g) | RRG (%) | Yield (g) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||
| Deuraechan | 3 | 26.3a |
1.74a | 42a | 166a | 14a | 86a | 6.1a | 28a | 80a | 2.9a | 1.9ab | 0.9bc | 69.1a | 32.4a | 9c | 25.0a | 87.3ab | 535bc |
| Boramchan | 3 | 21.0d | 1.53bc | 35bc | 129b | 11d | 60d | 5.7d | 24ab | 69ab | 2.8ab | 2.3a | 1.1a | 67.6a | 30.3b | 11a | 24.3a | 91.3a | 551b |
| a | 108 | 24.7ab | 1.63ab | 40ab | 156a | 13b | 79b | 6.0ab | 27a | 78a | 2.9a | 2.1ab | 1.0abc | 68.7a | 32.3a | 9bc | 24.2a | 84.1b | 520c |
| (20.1~27.1) | (1.29~1.88) | (26~51) | (107~197) | (11~15) | (65~93) | (5.7~6.3) | (15~38) | (41~118) | (2.6~3.1) | (1.3~3.1) | (0.6~1.6) | (62.3~74.3) | (28.3~35.7) | (8~13) | (21.7~27.2) | (74.1~93.0) | (477~554) | ||
| b | 42 | 22.6cd | 1.49c | 33c | 127b | 12c | 69c | 5.8c | 21b | 58b | 2.7b | 1.8b | 0.8c | 69.5a | 31.0ab | 11ab | 24.0a | 87.3ab | 467d |
| (19.2~26.5) | (1.32~1.77) | (24~44) | (97~172) | (10~14) | (57~85) | (5.6~6.1) | (13~31) | (35~90) | (2.5~3) | (1.2~2.6) | (0.5~1.3) | (63.5~75.0) | (26.9~33.5) | (9~12) | (21.0~27.1) | (69.4~94.9) | (410~501) | ||
| c | 13 | 23.7bc | 1.59bc | 40ab | 156a | 13bc | 75bc | 5.9bc | 28a | 81a | 2.9a | 2.2a | 1.1ab | 68.2a | 31.8ab | 10abc | 24.2a | 83.9c | 581a |
| (20.7~26.5) | (1.25~1.80) | (27~46) | (108~179) | (11~14) | (62~85) | (5.7~6.1) | (16~33) | (42~102) | (2.7~3.1) | (1.4~2.7) | (0.6~1.4) | (63.9~72.1) | (28.8~35.0) | (8~13) | (21.9~25.7) | (75.2~92.4) | (555~629) | ||
| Total | 163 | 24.1 | 1.59 | 38.2 | 149 | 13 | 76 | 5.9 | 26 | 73 | 2.8 | 2.0 | 1.0 | 68.8 | 31.9 | 10 | 24.1 | 84.9 | 512 |
| C.V.(%) |
163 | 8.3 | 8.2 | 15.4 | 15.1 | 8.7 | 10.3 | 3.4 | 20.0 | 23.0 | 3.6 | 15.0 | 20.0 | 3.5 | 5.0 | 11.3 | 4.5 | 5.3 | 7.2 |
zAbbreviations are listed in table
yMeans with same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
xC.V. = coefficient of variation
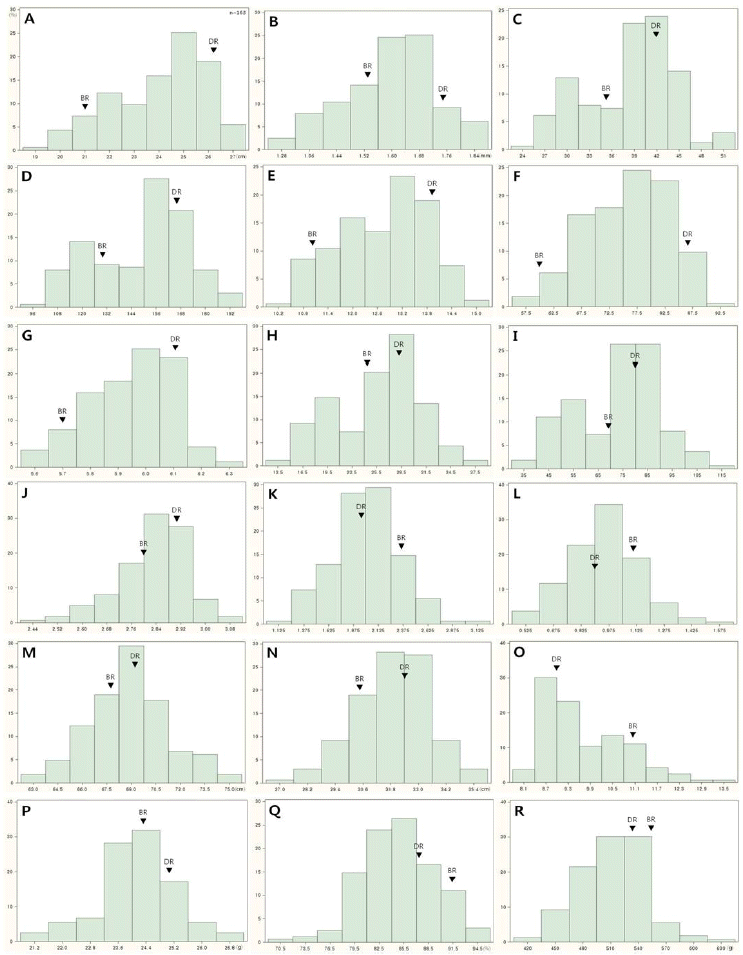
Panicle and yield-related traits of parents and selected DH lines at yield trial in 2013.
| Var./lines | PL |
TNN | NR | NS | NPr | NSPr | MNSPr | NSr | NSSr | MNSSr | Sr/Pr | SSr/SPr | CL (cm) | FL | PN | TGW (g) | RRG (%) | MRY (kg/10a) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||||||||
| Deuraechan (DR) | 26ab |
1.76ab | 32cde | 122bcd | 11abcd | 67bcd | 5.9bcde | 20de | 55efgh | 2.7ef | 1.7ef | 0.8ghi | 73ab | 36.0ab | 10efg | 25.3bc | 91.2a | 512b |
| Boramchan (BR) | 21e | 1.50def | 31cde | 111de | 9h | 51h | 5.5h | 22cde | 60def | 2.8cde | 2.3abc | 1.2bc | 71bcd | 32.1de | 13abc | 24.0ef | 91.4a | 567ab |
| HR28832-AC60-1-1 | 23cd | 1.62bcde | 31cde | 117bcde | 10fgh | 56gh | 5.8defg | 21cde | 61def | 2.8c | 2.2bcd | 1.1cd | 72bc | 34.2abcde | 12bcd | 23.7fg | 91.9a | 596a |
| (AC60) | ||||||||||||||||||
| HR28832-AC105-1-1 | 24cd | 1.69abc | 35abc | 132abc | 11bcde | 65cde | 5.8cdefg | 23bcd | 67bcde | 2.8c | 2.0de | 1.0def | 68e | 33.5bcde | 12abcd | 24.3e | 84.1bcd | 559ab |
| (AC105) | ||||||||||||||||||
| HR28832-AC107-1-1 | 26ab | 1.68abc | 39a | 149a | 12a | 73a | 6.0ab | 27ab | 75ab | 2.8cd | 2.2bcd | 1.0de | 73ab | 37.2a | 9h | 25.7ab | 83.8bcd | 516b |
| (AC107) | ||||||||||||||||||
| HR28832-AC109-1-1 | 23de | 1.34g | 26f | 101e | 10gh | 57g | 5.9abc | 16f | 44h | 2.7ef | 1.6fg | 0.7hi | 72bc | 31.0e | 13ab | 24.5de | 89.4ab | 561ab |
| (AC109) | ||||||||||||||||||
| HR28832-AC112-1-1 | 23de | 1.47efg | 31cde | 117bcde | 11def | 60gef | 5.7fg | 21de | 57efg | 2.7def | 1.9def | 0.9efg | 71bcd | 32.0de | 12cde | 23.2g | 92.2a | 549ab |
| (AC112) | ||||||||||||||||||
| HR28832-AC123-1-1 | 27a | 1.62bcde | 34abc | 133ab | 12abc | 71abc | 6.0a | 23cd | 62cdef | 2.8cdef | 1.9def | 0.9fgh | 73abc | 32.6cde | 10gh | 25.7ab | 85.3bcd | 541ab |
| (AC123) | ||||||||||||||||||
| HR28832-AC130-1-1 | 25ab | 1.66abc | 29ef | 115cde | 12abcd | 67bcd | 5.8cdef | 18ef | 47gh | 2.7ef | 1.5g | 0.7i | 73ab | 34.1abcde | 10fgh | 25.5abc | 87.6abc | 516b |
| (AC130) | ||||||||||||||||||
| HR28832-AC134-1-1 | 26ab | 1.58cdef | 37ab | 143a | 12ab | 72ab | 6.0a | 25abc | 71abcd | 2.8bc | 2.0cd | 1.0def | 75a | 35.8abc | 10gh | 25.0cd | 82.7cd | 500b |
| (AC134) | ||||||||||||||||||
| HR28832-AC149-1-1 | 23cd | 1.62bcde | 35abc | 132abc | 10efg | 58gf | 5.7g | 25abc | 74abc | 2.9ab | 2.4ab | 1.2ab | 68de | 31.3e | 11cdef | 23.3g | 85.0bcd | 564ab |
| (AC149) | ||||||||||||||||||
| HR28832-AC152-1-1 | 23cd | 1.65abcd | 38a | 148a | 11cde | 66cde | 6.0ab | 27ab | 82a | 3.0a | 2.4ab | 1.2ab | 72bcd | 35.9ab | 11cdef | 24.0ef | 89.5ab | 595a |
| (AC152) | ||||||||||||||||||
| HR28832-AC156-1-1 | 22de | 1.46fg | 29def | 110de | 10fgh | 56g | 5.7efg | 20def | 54fgh | 2.7cdef | 2.0de | 0.9defg | 71bcd | 35.5abc | 14a | 23.7fg | 90.9a | 590a |
| (AC156) | ||||||||||||||||||
| HR28832-AC161-1-1 | 25bc | 1.73abc | 38a | 146a | 11de | 63def | 5.9bcd | 28a | 83a | 3.0a | 2.6a | 1.3a | 69de | 35.1abcd | 11defg | 25.5abc | 84.9bcd | 605a |
| (AC161) | ||||||||||||||||||
| HR28832-AC178-1-1 | 25abc | 1.80a | 33bcde | 122bcd | 11cde | 63def | 5.8cdefg | 22cd | 58efg | 2.7f | 2.0de | 0.9defg | 70cde | 33.8bcde | 10gh | 26.1a | 80.7d | 507b |
| (AC178) | ||||||||||||||||||
| Mean | 24 | 1.61 | 33 | 125 | 11 | 62 | 5.8 | 62 | 22 | 2.8 | 2.0 | 1.0 | 71 | 34 | 11 | 24.6 | 87.5 | 552 |
| C.V. | 7.5 | 8.7 | 27.3 | 28.0 | 18.2 | 21.0 | 5.2 | 36.4 | 40.3 | 7.1 | 30.0 | 30.0 | 3.5 | 6.9 | 13.5 | 4.1 | 5.3 | 8.2 |
zAbbreviations are listed in table
yMeans with same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
Correlation coefficient among panicle-related traits and agronomic traits in 2013 (n=15).
| Traits | PL |
TNN | NR | NS | NPr | NSPr | MNSPr | NSr | NSSr | MNSSr | Sr/Pr | SSr/SPr | CL | FL | PN | TGW | RRG | MRY |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (x1) | (x2) | (x3) | (x4) | (x5) | (x6) | (x7) | (x8) | (x9) | (x10) | (x11) | (x12) | (x13) | (x14) | (x15) | (x16) | (x17) | (x18) | |
|
|
||||||||||||||||||
| x1 | 0.612* | 0.464ns | 0.558* | 0.902** | 0.907** | 0.728** | 0.279ns | 0.218ns | -0.008ns | -0.265ns | -0.213ns | 0.420ns | 0.473ns | -0.918** | 0.802** | -0.531* | -0.647** | |
| x2 | 0.568* | 0.564* | 0.584* | 0.519* | 0.209ns | 0.483ns | 0.443ns | 0.251ns | 0.197ns | 0.210ns | -0.233ns | 0.486ns | -0.684** | 0.571* | -0.457ns | -0.289ns | ||
| x3 | 0.984** | 0.581* | 0.582* | 0.423ns | 0.975** | 0.952** | 0.761** | 0.647** | 0.692** | -0.109ns | 0.523* | -0.570* | 0.232ns | -0.518* | 0.010ns | |||
| x4 | 0.671** | 0.687** | 0.560* | 0.932** | 0.914** | 0.739** | 0.543* | 0.588* | -0.023ns | 0.554* | -0.619* | 0.301ns | -0.532* | -0.040ns | ||||
| x5 | 0.987** | 0.713** | 0.384ns | 0.320ns | 0.059ns | -0.232ns | -0.182ns | 0.341ns | 0.549* | -0.882** | 0.660** | -0.548* | -0.650** | |||||
| x6 | 0.815** | 0.390ns | 0.334ns | 0.092ns | -0.232ns | -0.175ns | 0.393ns | 0.558* | -0.847** | 0.663** | -0.539* | -0.591* | ||||||
| x7 | 0.285ns | 0.272ns | 0.164ns | -0.212ns | -0.143ns | 0.450ns | 0.463ns | -0.536* | 0.541* | -0.375ns | -0.247ns | |||||||
| x8 | 0.992** | 0.847** | 0.798** | 0.835** | -0.217ns | 0.442ns | -0.404ns | 0.082ns | -0.437ns | 0.190ns | ||||||||
| x9 | 0.907** | 0.835** | 0.861** | -0.249ns | 0.407ns | -0.330ns | 0.021ns | -0.390ns | 0.278ns | |||||||||
| x10 | 0.871** | 0.857** | -0.301ns | 0.244ns | -0.047ns | -0.211ns | -0.164ns | 0.540* | ||||||||||
| x11 | 0.992** | -0.471ns | 0.102ns | 0.108ns | -0.318ns | -0.103ns | 0.598* | |||||||||||
| x12 | -0.426ns | 0.157ns | 0.060ns | -0.269ns | -0.148ns | 0.563* | ||||||||||||
| x13 | 0.354ns | -0.328ns | 0.249ns | 0.187ns | -0.468ns | |||||||||||||
| x14 | -0.466ns | 0.380ns | -0.160ns | -0.158ns | ||||||||||||||
| x15 | -0.743** | 0.551* | 0.679** | |||||||||||||||
| x16 | -0.579* | -0.584* | ||||||||||||||||
| x17 | 0.409ns | |||||||||||||||||
zAbbreviations are listed in table
Eigenvectors for the principal components of traits and correlation coefficient between the traits and principal components.
| Trait | Eigenvector | Correlation coefficient | ||||
|---|---|---|---|---|---|---|
|
|
||||||
| Prin1 |
Prin2 | Prin3 | Prin1 | Prin2 | Prin3 | |
|
|
||||||
| PL | 0.275 | -0.230 | -0.011 | 0.789** | -0.551* | -0.014ns |
| TNN | 0.246 | -0.015 | -0.375 | 0.707** | -0.037ns | -0.451ns |
| NR | 0.313 | 0.172 | 0.030 | 0.898** | 0.412ns | 0.036ns |
| NS | 0.326 | 0.127 | 0.087 | 0.937** | 0.306ns | 0.105ns |
| NPr | 0.292 | -0.198 | 0.027 | 0.838** | -0.476ns | 0.032ns |
| NSPr | 0.293 | -0.195 | 0.108 | 0.842** | -0.468ns | 0.130ns |
| MNSPr | 0.228 | -0.150 | 0.342 | 0.653** | -0.359ns | 0.411ns |
| NSr | 0.275 | 0.249 | 0.027 | 0.788** | 0.598* | 0.032ns |
| NSSr | 0.260 | 0.274 | 0.053 | 0.745** | 0.659** | 0.064ns |
| MNSSr | 0.176 | 0.329 | 0.145 | 0.506ns | 0.791** | 0.174ns |
| Sr/Pr | 0.103 | 0.389 | -0.036 | 0.297ns | 0.933** | -0.043ns |
| SSr/SPr | 0.123 | 0.377 | -0.014 | 0.352ns | 0.906** | -0.016ns |
| CL | 0.040 | -0.245 | 0.569 | 0.114ns | -0.589* | 0.684** |
| FL | 0.221 | -0.034 | 0.313 | 0.635* | -0.082ns | 0.376ns |
| PN | -0.287 | 0.178 | 0.112 | -0.825** | 0.427ns | 0.134ns |
| TGW | 0.203 | -0.241 | -0.227 | 0.582* | -0.579* | -0.273ns |
| RRG | -0.225 | 0.044 | 0.422 | -0.645** | 0.105ns | 0.507ns |
| MRY | -0.109 | 0.331 | 0.181 | -0.312ns | 0.795** | 0.218ns |
| Eigen value | 8.24 | 5.77 | 1.45 | |||
| Contribution (%) | 45.8 | 32.0 | 0.08 | |||
| Cumulative contribution (%) | 45.8 | 77.8 | 85.8 | |||
zPrin1 and Prin2 indicate the first and second component, respectively
Yield-related traits of parents and selected DH lines at yield trial in 2014.
| Var./lines | Heading date (DAS |
CL (cm) | PL (cm) | PN | NS | RRG (%) | TGW (g) | RRY (kg/10a) | BRR (%) | BRY (kg/10a) | MRY (kg/10a) | Index of MRY (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||
| Deuraechan (DR) | 107ay | 69a | 24a | 9c | 142a | 91.1ab | 25.8a | 641b | 81.6b | 523b | 481b | 100 |
| Boramchan (BR) | 108a | 69a | 20c | 12a | 108b | 94.5a | 24.4b | 736a | 82.5ab | 607a | 558a | 116 |
| HR28832-AC60-1-1 | 108a | 66bc | 22b | 11b | 129ab | 91.0ab | 24.5b | 772a | 81.0b | 625a | 575a | 119 |
| (AC60) | ||||||||||||
| HR28832-AC152-1-1 | 108a | 68ab | 22b | 11b | 140a | 90.5ab | 24.7b | 744a | 81.6b | 607a | 558a | 116 |
| (AC152) | ||||||||||||
| HR28832-AC156-1-1 | 108a | 64c | 20c | 12a | 108b | 94.8a | 24.0c | 741a | 83.2a | 617a | 567a | 118 |
| (AC156) | ||||||||||||
| HR28832-AC161-1-1 | 106a | 65bc | 22b | 11b | 143a | 86.3b | 25.9a | 748a | 81.1b | 607a | 559a | 116 |
| (AC161) | ||||||||||||
zAbbreviations are listed in table
yMeans with same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
Percent plant regeneration is the number of plants regenerated over number of cali plated
Percent green is the number of green plants regenerated over total plants regenerated
Percent albino is the number of albino plants regenerated over total plants regenerated
Double haploid lines are harvested plants having fertile grain
Upper and lower diagonal indicate the value in 2011 and 2012, respectively
Abbreviations are listed in table
Prin1 and Prin2 indicate the first and second component, respectively
Abbreviations are listed in table 1
Abbreviations are listed in table
Means with same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
C.V. = coefficient of variation
Abbreviations are listed in table
Means with same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)
Abbreviations are listed in table
Prin1 and Prin2 indicate the first and second component, respectively
Abbreviations are listed in table
Means with same letter are not significantly different at p < 0.05 (ANOVA followed by DMRT)