Abstract
The amylose contents of rice determine eating quality which is one of the major traits in rice breeding program. To identify the low-amylose gene of the japonica rice cultivar Baegjinju, genetic analysis was conducted using 200 F2 population derived from a cross between the japonica cultivars, Saeilmi and Baegjinju. Individual F2 plants were classified as wild type (translucent grain) and mutant type (dull grain) based on the grain appearance of brown rice. Two hundred F2 plants were segregated into 155 wild type plants and 45 mutant type plants, which fit the 3:1 ratio (x2 = 0.667, df = 1, p = 0.414) and this result indicated the low-amylose gene of Baegjinju is a single recessive gene which controls the amylose contents. Linkage analysis was conducted to localize the low-amylose gene of Baegjinju and fine mapped within an 800-kb interval between 17.5 to 18.8Mb on short arm of chromosome 10. Co-segregated SSR marker, RM25648 was developed and it could be useful for marker-assisted selection and determination of the genetic resource related with amylose contents in rice breeding.
-
Keywords: rice; low amylose; Baegjinju; molecular map; MAS.
서 언
우리나라는 국민 식생활의 서구화와 먹거리의 다양화 등으로 밥쌀용 쌀 소비가 지속적으로 감소하고 있으며 간단한 편이식품 에 대한 선호가 높은 1인 가구와 맞벌이 가구의 증가에 따라 쌀 가공식품에 대한 수요는 증가하는 있는 추세이다.
식량안보와 쌀 자급을 안정적으로 유지하기 위해서는 쌀 품질 의 고급화와 용도의 다양화를 통해 소비를 촉진하고 새로운 부가가치 창출하는 노력이 절실히 필요한 실정이다. 새로운 부가 가치 창출을 위해 기존의 밥쌀용 품종과 차별화된 특성을 가진 품종 개발이 지속적으로 추진되어 왔다. 그 예로 아밀로스 함량이 메벼와 찰벼의 중간특성을 가진 저아밀로스 품종 ‘백진주’가 개발되었으며 (
Hong et al. 2012a), 이후 ‘백진주1호’와 ‘설백’이 육성되었다(
Hong et al. 2012b,
Shin et al. 2012). 이러한 저아밀 로스 품종은 배유의 외관특성이 반투명(translucent) 또는 흐릿 한 흰색(hazy white)으로 나타나고, 아밀로스 함량은 7.1~14% 정도로 메벼와 찰벼의 중간 정도이다. 또한 메벼보다 전분의 점성이 강하고 팽화성이 높은 이화학적 특성을 가지고 있다(
Kim et al. 1999).
저아밀로스 형질에 대한 유전연구는 네팔 품종인 ‘Pokareli Mashino’을 이용해서 처음으로 불투명한 배유(opaque endosperm)에 대한 유전분석을 수행되었으며 waxy 유전자와 비슷한 배유 특성을 가지고 있고 대립유전자 관계에 있다고 보고되었다 (
Heu 1986). 또한, ‘고시히카리’의 돌연변이체인 ‘Milky Queen’과 ‘Kirara397’ 돌연변이체 ‘Hokkai287’에서 각각
Wx-mq와
Wx1-1이 밝혀졌다 (
Ando et al. 2010,
Sato et al. 2002). 이외에도 아밀로스 함량에 대한 돌연변이체들 중에서 waxy 유전자와 비대립관계에 있으면서 단순열성유전자를 가진 변이체가 탐색되었다 (
Okuno et al. 1983). 현재까지 보고된 유전자는 ‘Shiokaori’와 ‘Nihonmasari’에서 발생된 저아밀로스 변이체
lam(t)과
du(t)가 있고 (
Kinoshita & Kikuchi 1987), 자포니카 품종 ‘Kinmaze’에 MNU(N-methyl-N-nitrosourea) 를 처리하여 유기된 돌연변이체들에 대해 각각 유전분석을 수행 하였으며, 그에 따라
du-1, du-2, du-3, du-4 및
du-5의 유전자가 밝혀졌다 (
Satou & Omura 1981,
Yano et al. 1988). 국내에서는
Koh et al. (1997)에 의해 화청 유래의 du6A, du6B 와 Du7 유전자가 보고된 바 있다.
저아밀로스 유전자를 가진 품종은 등숙온도가 낮으면 현미의 외관특성이 맑고 투명하여 일반적인 메벼 품종과 구별하기 어렵 고, 이에 따라 교배 후대집단에서 표현형만으로는 선발이 어려운 경우가 많다. 또한 아밀로스 감소효과가 낮은 유전자를 모본으로 활용할 경우 저아밀로스 유전자의 효과가 불분명해 질 수 있고, 육종초기 세대에서는 분리계통이 혼재되어 있기 때문에 화학적 분석법인 아밀로스 함량 측정만으로는 초기세대 선발이 까다로 운 경우가 많다. 따라서 저아밀로스 유전자의 정밀한 유전자 지도를 작성하여 개발된 DNA마커를 이용할 필요가 있는데 이미 DNA마커를 이용한 교잡후대 계통의 선발에 이용한 사례가 보고된 바 있다 (
Ando 2008). 따라서, 본 연구는 배유의 전분다양 화 등 품질육종 분야에서도 분자육종체계를 확립하고자 국내에 서 개발된 ‘백진주’의 저아밀로스 형질에 대한 유전분석 및 분자 마커를 개발을 수행하였다.
재료 및 방법
유전분석용 재료 양성
본 연구를 수행하기 위하여 고품질 쌀인 ‘새일미’와 저아밀로 스 품종인 ‘백진주’를 교배하여 F2집단을 육성하였다. ‘백진주’ 는 고품질 품종인 ‘일품벼’의 수정란에 MNU를 처리하여 선발된 돌연변이 품종으로, 아밀로스 함량이 9.1%이며 쌀의 외관이 찹쌀과 멥쌀의 중간적 특성을 가진 뿌연 불투명한 배유를 가지고 있다(
Hong et al. 2012a). 모본으로 사용된 ‘새일미’는 쌀의 외관이 맑고 투명하면서 심복백이 없는 고품질 품종으로 불투명 한 저아밀로스 배유특성과 대조적인 특성을 가지고 있다 (
Lee et al. 2015). 모본 ‘새일미’와 부본 ‘백진주’ 각각 20주 및 F
1 식물체 15주를 국립식량과학원 남부작물부 벼 육종포장에서 30 × 15cm의 재식거리로 주당 1본으로 이앙하고, 시비량은 N-P
2O
5-K
2O=11-4.5-5.7 kg/10a으로 농촌진흥청 표준재배법 에 준하여 재배하였다.
배유의 표현형은 육안으로 현미 외관특성을 관찰하는 달관평 가방법에 따라 수행하였다 (
Koh et al. 1997). 항온기에서 1일간 건조된 현미 200립을 현미 색택에 따라 맑고 투명한 배유를 멥쌀의 translucent 형, 저아밀로스의 불투명한 배유를 dull 형으 로 분류하였다. 카이스퀘어(
X2 –test)검정을 통하여 적합도 분석 을 수행하였다.
현미를 Petri dish에서 발아시켜 생육이 양호한 개체를 육묘상 자에 이식하였다. 5엽기까지 정상적으로 생육한 식물체에서 유 전분석용 DNA 샘플을 채취하였다. Genomic DNA 추출은
Chen & Ronald (1999)의 Modified-CTAB 추출법을 이용하였 다. DNA 증폭은 Gene Amp. PCR 9700 (Applied Biosystems Co.)을 이용하여 10 ng의 genomic DNA, 0.2 mM dNTP mixture, 1x PCR buffer, 0.1 units Taq DNA polymerase (Genet-Bio Co., G-1002)의 농도로 반응액을 조성하여, denaturation을 95°C에서 30초, annealing 을 55°C에서 30초, polymerization을 72°C에서 1분으로 40cycle을 실시하였다. PCR 반응을 끝낸 증폭 산물은 4% 아크릴아마이드겔상에서 전기영동을 실시하여 GelStar (Lonza Co) 10배 희석액 50ml을 도포하여 10분간 상온에서 염색을 하였다. 염색을 마친 gel 염색 한 다음 UV transilluminator하에서 Gel Doc 시스템으로 촬영하 여 유전자형을 조사하였다.
‘백진주’의 저아밀로스 유전자 지도 작성 및 연관마커 탐색을 위해 모본인 ‘새일미’와 부본인 ‘백진주’의 다형성을 분석하였으 며, 총 110개를 SSR 마커를 사용하였다. 1차 연관분석에서는 저아밀로스 유전자가 위치한 연관 염색체 탐색하기 위해 새일미/ 백진주 F2 집단 200개체 중에서 표현형이 명확한 translucent형 과 dull 형을 각각 10개체 선발하였으며, 모부본 간에 다형성을 보인 마커와 조환가 분석을 수행하였다. 2차 연관분석에서는 새일미/백진주의 F2 집단 291개체를 이용하여 1차 연관분석에서 탐색된 SSR마커 및 인근영역에 존재하는 SSR 마커를 이용하여 연관분석을 수행하였다. 저아밀로스 유전자에 대한 물리적 유전 자 지도 작성은 ‘Nipponbare’유전자 지도를 기반으로 작성하였 다.
결과 및 고찰
저아밀로스 유전분석
‘백진주’의 저아밀로스 특성에 관한 유전분석을 수행하기 위 해 저아밀로스의 불투명한 배유가 명확하게 구분될 수 있도록 새일미/백진주 후대 F
2 종자를 30°C 항온기에 24시간 건조하여 수분함량을 14%이하로 낮추었다. 새일미/백진주의 F
2집단에서 translucent형과 dull 형은 각각 155개, 45개로서 3:1으로 분리하 였으며 (
X2 =0.667 p=0.414), 1개의 열성유전자에 의해 지배됨 을 확인하였다 (Table
1). 이 결과로 미루어 ‘백진주’의 유전양식 이 기존의 밝혀진 dull 유전자들과 같이 열성유전이라는 것을 확인할 수 있었다 (
Yano et al. 1988).
Kiswara et al. (2014)은 ‘백진주’와 저아밀로스 유전자 du12(t)를 가진 ‘밀양262호’가 교배된 후대집단에서 대립성검정을 수행하였고 translucent형과 dull형이 9:7로 분리되어 2개의 열성유전자에 의해 지배된다고 보고하였다. 이는 ‘백진주’의 저아밀로스 유전자와 ‘밀양262’호 의 du12(t)가 외관적으로 나타나는 현미의 색택에 미치는 영향은 비슷한 것으로 나타났으나 유전 분석을 통해 서로 다른 유전자라 는 것을 확인하였다. (Fig.
1).
Table 1.Genetic analysis of endosperm characters in F2 population derived from Saeilmi and Baegjinju.
Table 1.
|
Materials |
Generation |
No. of plants
|
Sum. |
Expected ratio |
X2
|
P |
|
translucent |
dull |
|
|
Saeilmi |
P1
|
20 |
|
20 |
- |
|
|
|
Baegjinju |
P2
|
|
20 |
20 |
- |
|
|
|
Saeilmi/Baegjinju |
F1
|
15 |
|
15 |
- |
|
|
|
F2
|
145 |
55 |
200 |
3 : 1 |
0.667 |
0.414 |
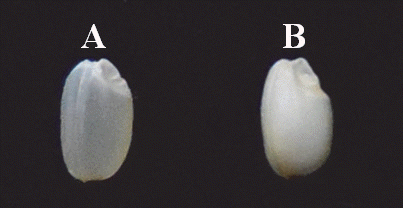
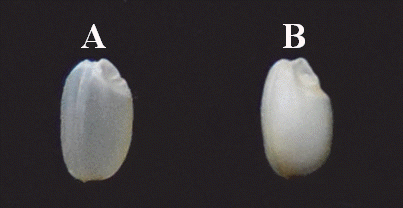
Fig. 1.Endosperm characteristics of parental varieties. (A) Saeilmi (translucent type), (B) Baegjinju (dull type).
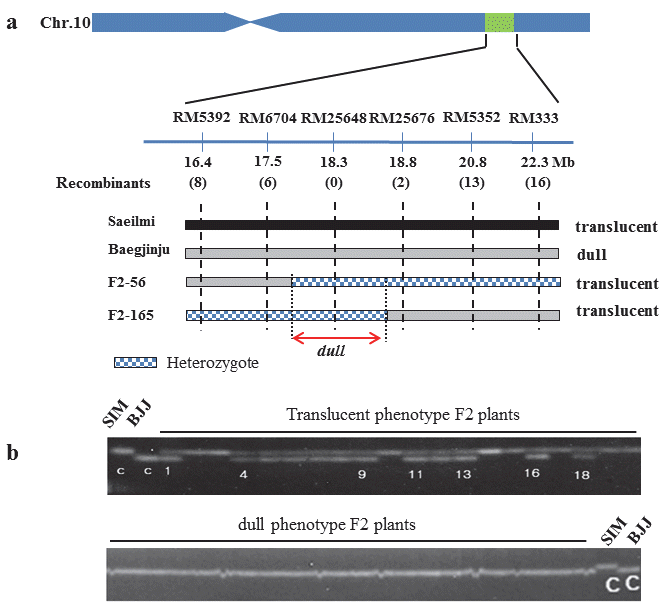
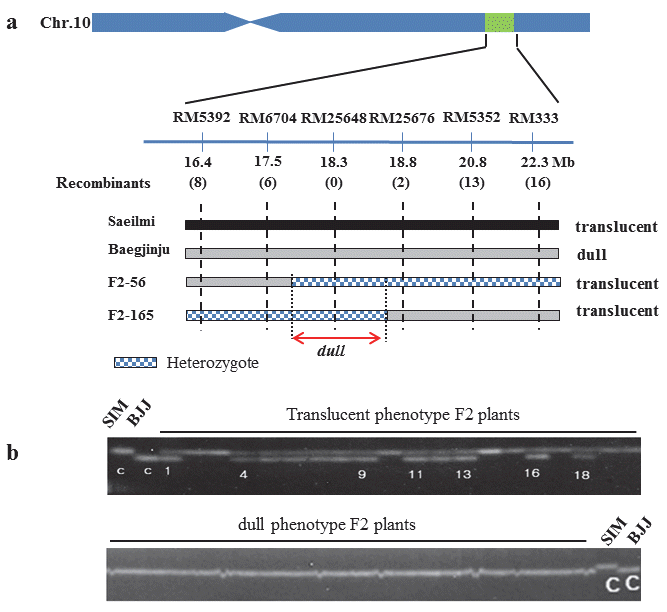
저아밀로스 연관분석
‘백진주’의 저아밀로스 관련 유전자 지도 작성 및 연관마커 탐색을 위해 모부본으로 사용된 ‘새일미’와 ‘백진주’에 대해 총 110개를 SSR 마커를 사용하여 다형성을 분석한 결과 RM545 등 24개의 다형성을 보이는 SSR 마커가 탐색되었다. 표현형이 명확한 translucent형과 dull형 F
2식물체를 각각 10개체씩 선발 하였으며, 모본과 부본에서 다형성을 보인 24개 SSR 마커를 이용하여 유전자형을 분석하였다. 24개 SSR마커 중 10번 염색 체상에 존재하는 RM6704에서 표현형과 유전자형이 일치하지 않는 재조환개체가 1개 탐색되었으며, 24개 마커 중에서 연관성 이 가장 높았다 (Fig.
2). 반면, 3번 염색체에 존재하는 RM545는 재조환 개체가 13개로 조환가가 50%이상 이었다. 일반적으로 질적형질에 대한 초기 연관마커 탐색을 위해 BSA (bulked segregant analysis) 방법 (
Mecheolmore et al. 1991)이 많이 사용되고 있어 본 연구에서도 BSA 방법을 적용하였으나, 연관 된 SSR마커를 찾지 못하였다 (data not shown). 이는 다형성 보인 마커들 중에서 가장 연관이 높았던 RM6704에서도 재조환 개체가 존재하였기 때문이며 모본과 부본이 국내육성 자포니카 품종으로 유전적 배경이 유사하고, 다형성 마커가 부족하여 BSA 방법으로 초기에 연관된 마커가 탐색되지 않은 것으로 판단된다.
Fig. 2.Genotype of linked marker RM6704 on chromosome10 and unlinked marker RM545 on chromosome 3 from pooled DNA sample. SIM; Saeilmi, BJJ; Baegjinju

벼 10번 염색체 17.5Mb에 위치한 RM6704 인근의 SSR 마커 를 제작하여 추가적으로 다형성을 분석하였으며, 16.8~22.3Mb 사이에 존재하는 RM5392등 6개의 다형성을 보인 마커를 선발 하였다. 새일미/백진주의 F
2 집단 291계통을 이용하여 표현형과 유전자형에 대해 연관분석을 수행한 결과 (Table
2), RM6704는 표현형과 유전자형이 다른 8개의 F
2개체가 탐색되었으며, 조환 가는 2.75%이였다. 반면, RM25648은 표현형과 유전자형이 일치하였으며, 조환가가 0.00으로 백진주의 저아밀로스 유전자 와 완전 연관된 마커일 가능성이 있다고 예상했다.
Table 2.Linkage analysis of the low amylose gene locus and SSR markers on chromosome 10 in the F2 population derived from a cross between Saeilmi and Baegjinju.
Table 2.
|
A |
B |
Segregation mode
|
Recombination ratio (%) |
Physical location (Mb) |
|
AAB_ |
AAbb |
AaB_ |
Aabb |
aaB_ |
aabb |
|
|
RM5392-dull |
86 |
0 |
144 |
7 |
11 |
43 |
6.19 |
16.8 |
|
RM25560-dull |
89 |
0 |
145 |
2 |
7 |
48 |
3.09 |
17.2 |
|
RM6704-dull |
88 |
0 |
147 |
2 |
6 |
48 |
2.75 |
17.5 |
|
RM25648-dull |
89 |
0 |
152 |
0 |
0 |
50 |
0.00 |
18.8 |
|
RM5352-dull |
81 |
1 |
147 |
17 |
13 |
32 |
10.65 |
20.8 |
|
RM333-dull |
84 |
1 |
141 |
20 |
16 |
29 |
12.71 |
22.3 |
유전자지도 작성 및 분자마커 선발
정밀 유전자 지도 작성을 위해 RM6704와 RM5352 사이 다형성이 보이는 SSR마커 RM25676를 추가적으로 선발하였 다. 목적한 저아밀로스 유전자와 거리를 좁히고자 RM6704와 RM5352에서 재조환이 일어난 F
2 식물체 각각 6개체와 14개체 를 선발하였다 (Fig.
3A). RM6704에서는 6개의 재조환 개체가 발생하였으며, RM25676에서는 2개의 재조환이 발생하였다. 또한, RM25648은 표현형과 유전자형이 일치하였다. 따라서, 본 연구에서 ‘백진주’의 저아밀로스 유전자는 10번 염색체의 장완에 RM6704와 RM25676사이에 존재하며, 물리적으로 17.5Mb ~ 18.8Mb 사이에 위치하였다. 또한 ‘백진주’의 저아밀 로스 유전자와 가장 밀접히 연관된 SSR marker RM25648를 선발하였으며 (Fig.
3B), 금후 MAS를 이용한 ‘백진주’의 저아밀 로스 유전자 선발에 활용될 것으로 기대된다.
Fig. 3.High-resolution linkage map of the dull region produced with the F2 population (a). The number of recombinant between the adjacent markers is indicated under the linkage map. Progeny testing of heterozygous recombinant delimited the dull locus to the region between SSR markers RM6704 and RM25676. Grey bars show the homozygous regions for ‘Baegjinju’ alleles; black bars indicate the homozygous regions for ‘Saeilmi’ alleles. Dotted bars represent the heterozygous region. (b) Co-segregation with the phenotype was analyzed by PCR amplification using the SSR marker RM25648 linked to the dull gene of Baegjinju. Saeilmi; SIM, Baegjinju ; BJJ
저아밀로스 유전자에 대한 분자생물학적 연구를 살펴보면, 찰성 유전자와 대립관계에 있는 ‘Milky Queen’의 Wx-mq와 ‘Hokkai287’의 Wx1-1에 관여하는 유전자는 GBSSI의 염기서 열 결실이 원인이라고 보고되었다 (Sato et al. 2002). 그리고 지금까지 밝혀진 12개의 dull 유전자들 중에서 du1, du2, du3 du12(t)에 대해 분자유전자 지도가 작성되었으며, 이들 위치는 각각 벼 염색체 2번, 3번, 6번, 10번에 위치하는 것으로 밝혀져 있다(
Kiswara et al. 2014,
Isshiki et al. 2008,
Zeng et al 2007). du-1과 du-2는 waxy pre-mRNA 유전자의 변이에 의해서 유전 자의 기능이 발현된다고 보고된 바 있다 (
Isshiki et al. 2008,
Zeng et al. 2007).
‘백진주’의 저아밀로스 유전자는 10번 염색체의 17.5Mb ~ 18.8Mb 사이에 존재하며, 이 영역은
Zeng et al. (2007)이 보고한 du1 유전자가 포함되어 있는데, waxy pre-mRNA 유전자의 SNP (single nucleotide polymorphism)변이가 원인인 것으로 밝혀져 있다. du1 유전자 특이적인 dCAPS marker를 이용하여 ‘백진주’의 dull 유전자를 분석한 결과
Zeng et al. (2007)이 보고한 du1 유전자와 동일한 SNP를 가지고 있음을 확인하였다 (data not shown).
Zeng et al. (2007)이 사용한 ‘Xiushui 11’ 돌연변이 계통 kl704와 실험재료는 다르지만 ‘백진주’의 저아밀 로스 유전자는 du1 유전자와 동일한 유전자인 것으로 판단된다. 본 연구의 내용이 유전자 기능분석 측면 신규성은 미흡하지만, 나고야의정서 발효 이후 유전자원에 대한 이익공유제의 현실화 및 유전자 기능에 대한 지식재산권 강화 등을 고려한다면 국내 육성품종의 유전자 기능에 관한 보고도 필요한 것으로 판단된다.
아밀로스는 가공적성이나 식미에 강하게 영향을 미치는 요소 이기 때문에 성분함량에 따라 다양한 용도로 활용되고 있다. 저아밀로스 품종의 전분은 점성이 강하고 팽화성이 크기 때문에 제과용이나 스낵식품 원료로 활용 가능성이 크다고 하였다(
Kim et al. 1999,
Hong et al. 2012b).
Kiswara et al.(2014)은 국내육 성 저아밀로스 품종인 ‘밀양262호’(건양2호)와 ‘백진주’ 및 ‘백 옥찰’과 대립성 검정을 통해 서로 독립적으로 유전하며, 이들 품종들이 가진 전분생합성 유전자들의 집적이 가능하다고 하였 다. 전분 생합성에 관여하는 다양한 유전자의 조합을 통해 쌀의 용도다양화 및 신형질 벼 품종 개발이 가능하며, 이를 위해서는 DNA마커를 이용한 선발기술이 효율적으로 활용 될 것으로 기대된다.
적 요
쌀을 식량생산 위주의 1차 산업으로만 간주하던 시각을 가공, 소비, 연관 산업을 포함하는 통합적인 관점에서 인식의 전환 및 새로운 가치 창출이 절실히 요구되고 있다. 쌀에서 다양한 전분특성을 가진 품종 육성을 위해서 ‘백진주’의 dull 유전자에 대한 유전학적인 연구를 수행하였다. ‘새일미’와 ‘백진주’의 F2 집단에서 dull형과 translucent형은 각각 62개, 218개로서 1 : 3으로 분리하였으며 1개의 열성유전자에 의해 유전됨을 확인하 였다. ‘백진주’의 저아밀로스 유전자는 10번 염색체의 장완에 RM6704와 RM25676사이에 존재하며, 물리적인 위치는 17.5Mb ~ 18.8Mb 사이에 존재하였다. 본 연구에서는 ‘백진주’ 의 저아밀로스 유전자와 가장 밀접히 연관된 SSR marker RM25648를 선발하였으며, 금후 MAS를 이용한 ‘백진주’의 저아밀로스 유전자 선발에 활용될 것으로 기대된다.
사 사
본 성과물(논문)은 농촌진흥청 연구사업(세부과제번호 : PJ010268012016)의 지원에 의해 이루어진 것임.
References
- 1. Ando I. Genetic analysis and breeding for low amylose content of rice. Gamma Field Symposia 2008. 47: 27-31.
- 2. Ando I, Sato H, Aoki N, Suzuki Y, Hirabayashi H, uroki M, Shimizu H, Ando T, Takeuchi Y. Genetic analysis of the low-amylose characteristics of rice cultivars Oborozuki and Hokkai-PL9. Breeding Science 2010. 60: 187-194.
- 3. Chen DH, Ronald PC. A rapid DNA minipreparation method suitable for AFLP and other PCR applications. Plant Mol Biol Rep 1999. 17: 53-57.
- 4. Heu MH. Inheritance of chalkiness of brown rice found in a non glutinous cultivar ‘Pokhareli Mashino’. Kor. J. Breed. Sci 1986. 18: 162-166.
- 5. Hong HC, Choi HC, Hwang HG, Kim YG, Moon HP, Kim HY, Yea JD, Shin YS, Choi YH, Cho YC, Baek MK, Lee JH, Yang CI, Jeong KH, Ahn SN, Yang SJ. A lodging-tolerance and dull rice cultivar Baegjinju. Kor. J. Breed. Sci 2012. 44: 51-56.
- 6. Hong HC, Yang SJ, Lee JH, Jung OY, Yang CJ, Choi YH, Kim YG, Lee KS, Choi IS, Cho YC, Kim MK, Lee JI, Jeong EK, Roh JH, Kim KJ, Hwang HG, Moon HP, Lee YT. A Medium-Maturing Dull cultivar Baegjinju 1. Kor. J. Breed. Sci 2012. 44: 57-61.
- 7. Isshiki M, Matsuda Y, Takasaki A, Wong HL, Satoh H, Shimamoto K. Du3, a mRNA cap-binding protein gene, regulates amylose content in Japonica rice seeds. Plant Biotechnol 2008. 25: 483-487.
- 8. Kim HK, Kim EY, Jeong YP. Physicochemical properties of non-glutinous, dull and glutinous rice grain in segregating populations of dull/glutinous crosses. Korean. J. Crop. Sci 1999. 44: 277-281.
- 9. Kinoshita T, Kikuchi H. Inheritance of amylose content in crosses of low amylose mutant RGN 4 1987. 83-85.
- 10. Kiswara G, Lee JH, Hur YJ, Cho JH, Lee JY, Kim SY, Sohn YB, Song YC, Nam MH, Yun BW, Kim KM. Genetic analysis and molecular mapping of low amylose gene du12(t) in rice (Oryza sativa L). Theor. Appl. Genet 2014. 27: 51-57.
- 11. Koh HJ, Cha KW, Heu MH. Inheritance and some physicochemical properties of newly induced low-amylose endosperm mutant in rice. Kor. J. Breed. Sci 1997. 29: 368-375.
- 12. Lee JH, Lee JY, Yoon YN, Kim SY, Hur YJ, Yeo US, Sohn YB, Song YC, Park DS, Nam MH, Cho JH. Enhancement of panicle blast resistance in Korean rice cultivar ‘Saeilmi’ by marker assisted backcross breeding. Plant Breed. Biotech 2015. 3: 1-10.
- 13. Michelmore RW, Paran I, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis a rapid method to detect markers in specific genomic regions by using segregating populations Proc. Natl. Acad. Sci 1991. 88(21):9828-9832.
- 14. Okuno K, Fuwa H, Yano M. A new mutant gene lowering amylose content in endosperm starch of rice Oryza sativa L. Japan. J. Breed 1983. 33: 387-394.
- 15. Sato H, Suzuki Y, Sakai M, Imbe T. Molecular Characterization of Wx-mq, a novel mutant gene for low-amylose content in endosperm of rice(Oryza sativa L). Breeding Science 2002. 52: 131-135.
- 16. Satoh H, Omura T. New endosperm mutationsinduced by chemical mutagens in rice, Oryza sativa L. Japan. J. Breed 1981. 31: 316-326.
- 17. Shin YS, Won YJ, Ahn EK, Kim YK, Kim MK, Hwang HG, Yoon YH, Lee JH, Choi IS, Yoon KS, Oh SK, Kang KH, Shon JY, Jeon YH, Shin JC. An early maturing, blast resistant and low amylose rice variety Seolbaek. Kor. J. Breed. Sci 2012. 44: 597-601.
- 18. Yano M, Okuno K, Satoh H, Omura T. Chromosomal location of genes conditioning low amylose content of endosperm starches in rice Oryza sativa L. Theor. Appl. Genet 1988. 76: 183-189.
- 19. Zeng D, Yan M, Wang Y, Liu X, Qian Q, Li J. Du1, encoding a novel Prp1 protein, regulates starch biosynthesis through affecting the splicing of Wxbpre-mRNAs in rice (Oryza sativa L). Plant Mol. Biol 2007. 65: 501-509.
Citations
Citations to this article as recorded by