적요
The 12 cultivars of the Jeju native Citrus are considered to have originated from China. However, the origin of the cultivar ‘Byungkyool’ (Citrus platymamma Hort. ex Tanaka) is not clearly known. We performed PCR analysis by using three primer sets designed from the internal transcribed spacer (ITS) region of nuclear ribosomal DNA (nrDNA) to analyze the phylogenetic relationship between the traditional citrus cultivars and the Byungkyool cultivar. Sequence length of the nrDNA ITS1 region of JNCPCRI (Jeju Native Citrus platymamma Citrus Research Institute) cultivar was 247 bp, 8the ITS2 region was 228 bp and the total ITS region (ITS1-5.8S-ITS2) was 638 bp. Analysis of the genetic relationship based on the sequence analysis at the ITS region of the JNCPCRI cultivar revealed that the ITS1 region of the cultivar was genetically the same as that of the Byungkyool (JQ990189) cultivar, and the ITS2 region was genetically similar to the Binkyool (JQ990180), Hongkyool (JQ990178), Dangyooja (JQ990179), and Pyunkyool (JQ990181) cultivars. Moreover, the total ITS region in the 5.8S rDNA region was genetically similar to the Hongkyool (JQ990178) cultivar. In addition, the total ITS region of the JNCPCRI cultivar was the most closely related to the Cheongkyool (JQ990183) cultivar and has been reported to originate from the Binkyool (JQ990180) and Pyunkyool (JQ990181) cultivars. Although the JNCPCRI cultivar was morphologically the same as the Byungkyool (JQ990189) cultivar, the ITS region showed genetic heterogeneity. Taken together, we conclude that the genetic variation in the ITS region of JNCPCRI cultivar suggests that it was propagated through fertilization with the surrounding citrus cultivars.
서 언
제주도내 재배되었던 재래감귤 품종들로는 금귤, 감자, 당감 자, 산귤, 청귤, 동정귤, 유감, 당유자, 홍귤, 유자, 당금귤, 석금 귤, 왜귤, 편귤, 사두감, 주감, 광귤, 빈귤, 지각, 병귤, 진귤 등 22개 품종들이 알려지고 있으나(
Kim et al. 2001), 이 재래감 귤 품종 중 생식용으로는 맛이 없고 품질이 떨어지는 품종들은 차차 새로운 품종으로 개량되어 지금은 병귤, 당유자, 유자, 청귤, 동정귤, 홍귤, 진귤 등 12가지 품종만이 보존되고 있다 (
Kim et al. 2001). 고문서에 따르면 이 재래감귤 품종들은 선사시대부터 제주도에서 자생한 것으로 추정되며, 분명한 기 원에 대해서는 불명확 한 상태이다(
Jung et al. 2005). 최근, 미국 식물 유전자원 시스템(U.S. National Plant Germplasm System)에 따르면(
USDARS 2016), 이들 대부분의 품종들은 중국 혹은 일본 등 아시아 지역에 분포되어 있음을 알 수 있으나, 병귤 품종에 대한 정확한 분류학적 연구라든지 기원 연구는 미흡한 실정이다.
감귤에서 계통분류학적 연구는 형태학적(
Barrett & Rhodes 1976,
Scora 1975), 생화학적(
Hirai et al. 1986,
Herrero et al. 1996) 그리고 isozyme(
Herrero et al. 1996), restriction fragment length polymorphism(RFLP)(
Federici et al. 1998), random amplified polymorphic DNA(RAPD)(
Deng et al. 1995), inter simple sequence repeat(ISSR)(
Gulsen & Roose 2001), simple sequence repeat(SSR)(
Luro et al. 2008) 등과 같은 분자마커를 이용하여 주로 연구되었다. 이들 분자마커 기술 은 많은 식물들의 분류학적 연구에 광범위하게 이용되고 있다 (
Hazarika et al. 2014,
Jung et al. 2005,
Tingey & Tufo 1993,
Whitkus et al. 1994). 또한, 세포질내에 존재하는 엽록체 및 미토콘드리아 게놈의 유전자 및 유전자간 염기서열 변이를 활용 한 연구가 이용되고 있다(
Cheng et al. 2005,
Deng et al. 2007,
Froelicher et al. 2011,
Jung et al. 2005,
Olmstead & Palmer 1994,
Penjor et al. 2013). 최근에는 세포질내에 존재하는 18S, 5.8S, 28S rRNA를 암호화하는 종 보존부위인 핵 리보솜 DNA (nuclear ribosomal DNA, nrDNA)내 Internal Transcribed Spacer(ITS) 영역은 감귤류 분류군들에 대한 계통분류학적 연구 에 광범위하게 이용되고 있다(
Baldwin et al. 1995,
Hynniewta et al. 2014,
Pessina et al. 2011,
Sun et al. 2015,
Xu et al. 2006). ITS 영역은 핵 게놈에 존재하며 nrDNA영역의 일부로 반복적인 염기서열 형태로 구성되어 있으며 양친으로부터 유전 되는 특성을 갖고 있다. rRNA 영역은 근연종간에 거의 유전적 변이가 없는데 반하여 ITS 영역은 비교적 변이가 많이 일어남으 로 종속 구별에 있어서 유익하게 이용되고 있다.
제주감귤의 계통발생과 관련된 연구들로는 형태학적 특징 (
Kim 1988), isozymes(
Moon et al. 1992), RAPD 분석(
Oh 1996), 엽록체의 게놈을 이용한 분석을 통하여 연구되고 있다. 그러나 이들 분석 방법들은 돌연변이에 의한 품종들간을 구별하 기 힘들고(
Rahman et al. 2001), 재현성이 떨어진다는 단점을 갖고 있다(
Hazarika et al. 2014). 또한, 일부 교잡종들은 양친의 미토콘드리아가 유전된다는 점에서 모계유전과 관련된 엽록체 의 게놈을 이용한 분석 방법은 계통발생 분류에 있어 오류가 발생할 수 있다는 단점을 갖고 있다(
Froelicher et al. 2011). 따라서, 본 연구에서는 양친으로부터 유전되는 특성을 갖고 있는 ITS 영역의 염기서열을 이용하여 본 연구소 내 재배되고 있는 병귤 품종과 GenBank에 등록된 병귤 품종간의 유연관계 분석과 재래감귤 품종들 간의 계통분류학적 유연관계를 조사하였다.
재료 및 방법
식물 재료 및 전체 게놈 DNA 추출
본 연구에 사용된 제주도 재래감귤 12품종[진귤(Citrus sunki Hort. ex Tanaka), 지각(Citrus aurantium L.), 병귤(Citrus platymamma Hort. ex Tanaka), 동정귤(Citrus erythrosa Hort. et Tanaka), 홍귤(Citrus tachibana Tanaka), 청귤(Citrus nippokoreana Tanaka), 당유자(Citrus grandis Osbeck), 소유자 (Citrus junos Sieb. ex Tanaka), 사두감(Citrus pseudogulgul Hort. ex Tanaka), 감자(Citrus benikoji Hort. ex Tanaka), 빈귤 (Citrus leiocarpa Hort. ex Tanaka), 편귤(Citrus tangerina Hort. ex Tanaka)]의 잎들은 2015년 9월 14일 감귤 연구소 내 보존 재배되고 있는 품종들로부터 수집하였다. 재래감귤 12품종의 잎들에 대한 전체 게놈 DNA 추출은 Automatic Nuclear extraction(MX 16, Promega, Madison, WI, USA)을 이용하여 추출하였다. 추출한 식물 전체 게놈 DNA는 -70°C에 보관하여 실험에 사용하였다.
ITS 영역 염기서열 구성 및 프라이머 디자인
재래감귤 중 병귤 품종의 계통분류학적 연구를 위한 프라이머 디자인은 핵 게놈에 존재하며 핵 리보솜 DNA 영역을 구성하고 있는 모든 유전자들을 NCBI(National Center for Biotechnology Information)의 GenBank에 등록된 염기서열을 다운받아 이용하였다.
18S ribosomal RNA 영역의 유전자 염기서열은 손바닥 선인 장(Opuntia ficus-indica: Aceesion No AB250211), 기주밀감 (C. kinokuni: Aceesion No. JQ990159), 자몽(Citrus x paradis: Aceesion No. FJ641956), 감자(C. benikoji Hort. ex Tanaka: Aceesion No. JQ990186) 그리고 당유자(C. grandis Osbeck: Aceesion No. JQ990179)의 공통된 부분을 갖고 있는 18S ribosomal RNA 유전자부위로 구성하였고, ITS1, 5.8S ribosomal RNA, ITS2, 28S ribosomal RNA 유전자의 염기서 열 구성은 기주밀감(Aceesion No. JQ990159)의 유전자부위를 이용하였다.
ITS1 영역의 유전자를 증폭시키기 위하여, ITS1F1의 프라이 머는 18S rDNA와 ITS1 부위를 포함하도록 제작하였고 ITS1R1 의 프라이머는 ITS1와 5.8S rDNA 부위를 포함하도록 디자인 하였다. 구성된 프라이머 조합은 PCR에 사용하였다.
ITS 영역의 PCR 증폭 및 산물 정제
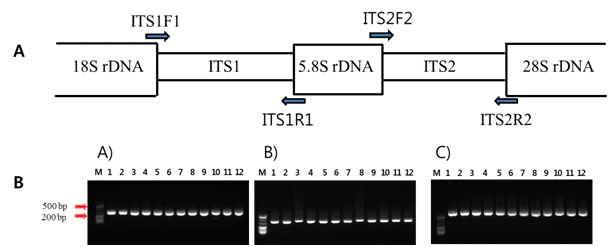
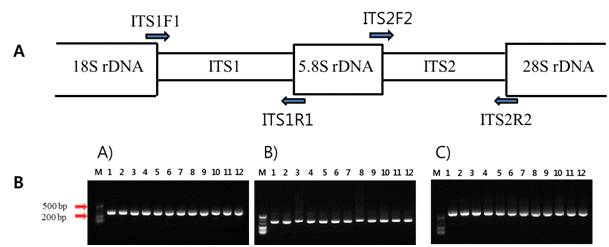
핵 ribosomal DNA의 ITS1 그리고 ITS2 부위에 대한 PCR 증폭 산물을 얻기 위하여, ITS1과 ITS2 부위를 포함하고 있는 ITS1F1-ITS1R1 그리고 ITS2F2-ITS2R2 프라이머 조합을 PCR 증폭에 사용하였다(Fig.
1A). 또한, ITS1부터 ITS2까지의 ITS 전체 영역(ITS1-5.8S-ITS2)을 PCR 증폭 시키기 위하여 ITS1F1와 ITS2R2 프라이머를 이용하였다(Fig.
1A). ITS영역 의 증폭을 위한 PCR 반응액은 AccuPower PCR Premix(Bioneer, Corp., Daejeon, Korea)[250μM dNTP, 1.5mM MgCl
2, 1.0unit
Taq DNA polymerase, 10mM Tris-HCl(pH 9.0), 40mM KCl]에 주형 DNA 25-50ng 그리고 0.5pmol의 프라이머를 첨가하여 20ul로 조정하여 사용하였다. ITS 영역의 PCR 반응은 Takara PCR Thermal cycle(Takara, Bio Inc., Japan)에서 35cycle를 실시하였으며, denaturation은 94°C에서 30초, annealing은 55°C에서 30초, 그리고 extension 을 72°C에서 40초간 수행하였다. 증폭된 PCR 생성물은 1.2% agarose gel에서100V로 30분 동안 전기영동 하여 나타나는 밴드로 확인하였다. Agarose gel에서 확인된 PCR 증폭 산물은 GgenAll
® Gel 정제 kit(GgenAll Biotechnology, Co., Seoul, Korea)를 이용하여 정제하였으며, 염기서열 결정은 ㈜ 솔젠트 (Solgent, Co., Daejeon, Korea)에 의뢰하였다.
Fig. 1.Nucleotide sequence of nrDNA for the primer design of ITS regions and the of JNCPCRI rDNA regions. A: Sequence of 18 S ribosomal RNA gene regions were constructed by of Opuntia ficus-indica (Acc. No AB250211), C. kinokuni (Acc. No. JQ990159) and Citrus x paradis (Acc. No. FJ641956). ITS1, 5.8S rRNA, ITS2 and 28S rRNA gene region were constructed by sequence of C. kinokuni (Acc. No. JQ990159). ITS1 and ITS2 spacer region gene was underlined. 18S, 5.8 and 28S ribosomal RNA gene regions were expressed by double arrow. ITS1F1, ITS1R1, ITS2F2 and ITS2R2 primers site were described by block box. B: JNCPCRI cultivar used in this study is ‘Byungkyool’ (C. platymamma Hort. ex Tanaka) harvested in Citrus Research Institute. ITS1 and ITS2 spacer region was underlined and ITS2 region was represented by the bold underline. 18S, 5.8 and 28S rDNA regions were expressed by double arrow.

ITS 영역의 sequence alignment 와 계통분석
본 연구소 내 재배되고 있는 재래감귤 병귤 품종에서 분리된 ITS 영역의 nucleotide (JNCPCRI)와 유전자 은행(GeneBank, NCBI)에 등록된(
Sun et al. 2015) 다른 재래감귤 11품종들 간의 계통분석을 위하여, 재래감귤 11품종의 염기는 유전자 은행 (GeneBank, NCBI)에서 다운받았다(Table
1). 모든 재래감귤의 염기서열은 fasta format으로 전환한 후 Bioedit(
Hall, 1999)을 이용하여 편집하였으며 no gap으로 정렬한 후 multi Clustal W로 정렬하여 저장하였다. 계통학적 분석은 MEGA 5.0 프로그 램을 사용하였고, 유전적 거리 계산은 kimura 2 - parameter를 이용하였으며. UPGMA기법으로 분계도를 작성하였다. 그리고, Bootstrap 값은 1,000회 반복하여 수행하였다.
Table 1.List of Jeju native Citrus cultivars used in this study and their relevant information of specimen voucher and NCBI accession number.
Table 1.
|
No. |
Cultivars
|
Tribe |
Specimen voucher |
GenBank acc. No. |
|
Generic name |
Scientific name |
|
|
1 |
Dangyooja |
C. grandis Osbeck |
Cephalocitrus
|
kk-70 |
JQ990179 |
|
2 |
Binkyool |
C. leiocarpa Hort. ex Tanaka |
Sinocitrus
|
kk-71 |
JQ990180 |
|
3 |
Pyunkyool |
C. tangerina Hort. ex Tanaka |
Sinocitrus
|
kk-72 |
JQ990181 |
|
4 |
Cheongkyool |
C. nippokoreana
|
Sinocitrus
|
kk-74 |
JQ990183 |
|
5 |
Jikak |
C. aurantium L. |
Sinocitrus
|
kk-75 |
JQ990184 |
|
6 |
Sadookam |
C. pseudogulgul Hort. ex Tanaka |
Sinocitrus
|
kk-76 |
JQ990185 |
|
7 |
Kamja |
C. benikoji Hort. ex Tanaka |
Sinocitrus
|
kk-77 |
JQ990186 |
|
8 |
Dongjeongkyool |
C. erythrosa Hort. et Tanaka |
Sinocitrus
|
kk-78 |
JQ990187 |
|
9 |
Jinkyool |
C. sunki Hort. ex Tanaka |
Sinocitrus
|
kk-79 |
JQ990188 |
|
10 |
Byungkyool |
C. platymamma Hort. ex Tanaka |
Sinocitrus
|
kk-80 |
JQ990189 |
|
11 |
Hongkyool |
C. tachibana Tanaka |
Sinocitrus
|
kk-69 |
JQ990178 |
결과 및 고찰
ITS 영역의 염기서열분석
본 연구는 재래감귤 12품종 중 JNCPCRI 병귤 품종에 대한 계통학적 유연관계분석을 위하여 ITS1, ITS2 그리고 전체 ITS 유전자 영역(ITS1-5.8S-ITS2)의 프라이머 조합을 이용하여 PCR를 수행하였다. 그 결과 Fig.
1에서 보는 것처럼 JNCPCRI 병귤을 포함한 재래감귤 12품종들의 ITS1과 ITS2 영역은 약 225-300bp의 PCR 증폭산물을 얻었다. 또한 전체 ITS 유전자 영역에 대한 JNCPCRI 병귤을 포함한 재래감귤 12품종 들의 PCR 결과는 약 550-700bp 의 증폭산물을 획득하였다(Fig.
2B-
C). 이 PCR 결과는
Sun et al. (2015)이 보고한 재래감귤 품종들의 ITS 영역에 대한 길이와 비슷하였다. 따라서, 이 프라이 머 조합은 재래감귤의 계통학적 유연관계 분석에 적용 할 수 있을 것으로 사료된다.
Fig. 2.PCR amplification using the nuclear internal transcribed spacer of Jeju Native Citrus. A: Ribosomal-RNA - coding gene regions (18 S, 5.8 S and 28 S) and spacer regions (ITS1 and ITS2) are indicated in the boxes. The arrows denote the positions of primers for PCR amplification. B: ITS PCR amplification of Jeju Native Citrus. A) PCR amplification of ITS1 region (ITS1F1 and ITS1R1 primer use); B) PCR amplification of ITS2 region (ITS2F2 and ITS2R2 primer use); C) PCR amplification of ITS1-5.8S rDNA-ITS2 region (ITS1F1 and ITS2R2 primer use). M; 20 bp Takara marker, lane 1; ‘Binkyool’, lane 2; ‘Cheongkyool’, lane 3; ‘Hongkyool’, lane 4; ‘Dongjeongkyool’, lane 5; ‘Kamja’, lane 6; Jinkyool’, lane 7; ‘Byungkyool’, lane 8; ‘Yuzu’, lane 9; ‘Pyunkyool’, lane 10; ‘Dangyooja’, lane 11; ‘Sadookam’ and lane 12; ‘Jikak’.
이들 프라이머 조합들(Fig.
2A)을 이용하여 본 연구소 내에서 재배되고 있는 재래감귤의 ITS1(18S, 5.8S rDNA 영역의 프라 이머 서열를 제외한) 염기서열 길이는 246-248bp, ITS2(5.8S, 28S rDNA 영역의 프라이머 영역을 제외한 염기서열의 길이)는 227-228bp였고, 전체 ITS(18S, 28S rDNA 영역의 프라이머 서열를 제외한)유전자 영역은 636-638bp였다(자료 미제시). 위 결과는 ITS 영역을 이용한
Sun et al. (2015)의 결과와 유사하였 으며, 병귤(JNCPCRI) 품종의 ITS1 영역 염기서열 정보는 Fig.
2B에서 보는 봐와 같이 247bp, 5.8S ribosomal RNA유전자의 염기서열 길이는 163bp 그리고 ITS2영역의 염기서열 길이는 228bp이었고, 전체 ITS 유전자 영역의 염기서열 길이는 638bp 였다. 또한, ITS1, 5.8S ribosomal RNA 그리고 ITS2 영역의 G(구아닌) 그리고 C(시토신)의 함량은 각각 70.85%, 54.60% 그리고 70.61%였다(Table
2). Table
2에서 보는 봐와 같이 이전 연구자(
Sun et al. 2015)의 병귤(JQ990189) 품종의 ITS1과 5.8S ribosomal RNA 유전자 영역의 염기서열 길이는 본 연구 결과와 같았다. 따라서 JNCPCRI병귤 품종의 ITS1 영역은 GenBank에 등록된 병귤(JQ990189)의 유전자와 거의 비슷한 크기를 가지고 있는 것으로 확인되었다. 그러나 JNCPCRI 병귤 품종의ITS2 영역 염기서열 길이는 228bp로 GenBank에 등록된 병귤 (JQ990189)의 염기서열보다는 길이가 길며 G(구아닌) 그리고 C(시토신)의 함량 또한 70.61%로 높았다. JNCPCRI 병귤 품종 의ITS2 영역 염기에 대하여 NCBI Blast로 상동성을 조사한 결과 빈귤(JQ990180), 편귤(JQ990181) 그리고 홍귤 (JQ990178) 품종들과는 90% 이상의 상동성을 보였으나, 병귤 (JQ990189) 품종과는 약61%로 나타나 큰 상동성을 보이지 않았다. 즉, 같은 제주도 지역 내 재배되고 있는 병귤 품종들이지 만 이들 품종들 간에 차이를 보이는 이유는 재래감귤 품종들의 번식자체가 수분 수정을 통한 종자번식으로 자생되어 왔기 때문 에
Sun et al. (2015)의 연구결과와 차이점을 보이는 것이라 생각된다.
Table 2.Sequence length and G+C content (%) of the JNCPCRI ITS gene region and comparison between JNCPCRI and ‘Byungkyool’ cultivar registered in the GenBank.
Table 2.
|
Cultivars |
Sequence length (bp)
|
G+C content (%)
|
|
ITS1 |
5.8S |
ITS2 |
ITS1 |
5.8S |
ITS2 |
|
|
JNCPCRIZ)
|
247 |
163 |
228 |
70.85 |
54.60 |
70.61 |
|
C. platymammaY)
|
247 |
163 |
227 |
70.85 |
54.60 |
70.48 |
ITS1 영역의 유전적 유연관계분석
JNCPCRI 병귤 품종과 GenBank에 등록된 재래감귤 11품종 의 ITS1 영역에 대한 염기서열 분기값(Sequence divergency)은 Kimura 2 - parameter 방법으로 계산하였다. 그 결과 JNCPCRI 병귤 품종은 GenBank에 등록된 병귤(JQ990189) 품종과의 염 기서열 분기값이 0로 다른 품종들에 비하여 염기서열의 다양성 정도가 가장 낮았으며, 염기서열이 동일한 것으로 확인되었다. 그 다음으로 홍귤(JQ990178) 품종이 0.00426로 병귤 (JQ990189) 품종 다음으로 염기서열 다양성 정도가 가장 낮았다 (Table
3). 또한, 빈귤(JQ990180), 청귤(JQ990183), 그리고 편 귤(JQ990181) 3품종이 모두 0.00852, 동정귤(JQ990187) 그리 고 지각(JQ990184) 2품종이 모두 0.00855, 당유자(JQ990179) 품종이 0.01282, 진귤(JQ990188) 품종이 0.02609 순으로 염기 서열 다양성 정도가 낮았다(Table
3). 그러나 사두감 (JQ990185), 감자(JQ990186) 품종의 염기서열 분기값은 각각 0.12647, 0.15205로 재래감귤 11품종 중 염기서열 다양성 정도 가 가장 높았다. 즉, 유전적으로 상이한 품종일 것이라고 생각된 다(Table
3). 또한, GenBank에 등록된 재래감귤 11품종의 ITS1 영역에 대한 염기들을 이용하여 JNCPCRI 병귤 품종간의 유연 관계는 MEGA 5.0 프로그램으로 분석하여 Group I과 Group II의 2개 그룹의 분류군으로 나누었다(Fig.
3A). Group I은 청귤 (JQ990183), 홍귤(JQ990178), 진귤(JQ990188), 지각 (JQ990184), 당유자(JQ990179), 편귤(JQ990181), 동정귤 (JQ990187), 빈귤(JQ990180), 병귤(JQ990189), 그리고 JNCPCRI 병귤 품종들이 포함되어 있으며, Group II에는 사두 감(JQ990185)과 감자(JQ990186) 품종으로 구성되어 있다(Fig.
3A). 또한, Group I은 빈귤(JQ990180), 편귤(JQ990181), 청귤 (JQ990183), 당유자(JQ990179)로 구성된 Ia, 홍귤(JQ990178), 병귤(JQ990189), JNCPCRI 품종으로 구성된 Ib, 지각 (JQ990184), 동정귤(JQ990187) 품종으로 구성된 Ic 그리고 단일로 구성된 Id 진귤(JQ990188) 품종의 4개 분계조로 다시 나누었다(Fig.
3A). ITS1영역의 염기를 이용하여 JNCPCRI 병귤 품종과의 유연관계를 조사한 결과 JNCPCRI 병귤 품종은 Group Ib속하며 GenBank에 등록된 병귤(JQ990189) 품종과는 100% 동일한 품종이며, 홍귤(JQ990178) 품종과는 형태학적으 로 서로 다르나 ITS1 영역에 100% 일치하는 결과를 얻었다.
Table 3.Sequence divergences in the ITS1 gene regions of JNCPCRI cultivar and related Jeju Native Citrus Varieties.
Table 3.
|
No. |
Cultivars |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
|
1 |
C. aurantium (JQ990184) |
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
C. benikoji (JQ990186) |
0.15205 |
|
|
|
|
|
|
|
|
|
|
|
|
3 |
C. erythrosa (JQ990187) |
0.00855 |
0.14118 |
|
|
|
|
|
|
|
|
|
|
|
4 |
C. leiocarpa (JQ990180) |
0.01718 |
0.16269 |
0.01718 |
|
|
|
|
|
|
|
|
|
|
5 |
C. grandis (JQ990179) |
0.02152 |
0.16784 |
0.02152 |
0.00425 |
|
|
|
|
|
|
|
|
|
6 |
C. nippokoreana (JQ990183) |
0.01718 |
0.16269 |
0.01718 |
0.00000 |
0.00425 |
|
|
|
|
|
|
|
|
7 |
C. platymamma (JQ990189) |
0.00855 |
0.15205 |
0.00855 |
0.00852 |
0.01282 |
0.00852 |
|
|
|
|
|
|
|
8 |
C. pseudogulgul (JQ990185) |
0.13054 |
0.05732 |
0.12013 |
0.14083 |
0.14585 |
0.14083 |
0.13054 |
|
|
|
|
|
|
9 |
C. sunki (JQ990188) |
0.02609 |
0.15205 |
0.01724 |
0.03496 |
0.03938 |
0.03496 |
0.02609 |
0.13054 |
|
|
|
|
|
10 |
C. tachibana (JQ990178) |
0.01288 |
0.15758 |
0.01288 |
0.00425 |
0.00853 |
0.00425 |
0.00426 |
0.13583 |
0.03058 |
|
|
|
|
11 |
C. tangerina (JQ990181) |
0.01718 |
0.16269 |
0.01718 |
0.00000 |
0.00425 |
0.00000 |
0.00852 |
0.14083 |
0.03496 |
0.00425 |
|
|
|
12 |
JNCPCRI |
0.00855 |
0.15205 |
0.00855 |
0.00852 |
0.01282 |
0.00852 |
0.00000 |
0.13054 |
0.02609 |
0.00426 |
0.00852 |
|
Fig. 3.Phylogenetic analysis of the nrDNA ITS region isolated in JNCPCRI and other Jeju Native Citrus Cultivars (registered in the GenBank). A: Phylogenetic analysis of the nrDNA ITS1 region, B: Phylogenetic analysis of the nrDNA ITS2 region, C: Phylogenetic analysis of the nrDNA total ITS region (ITS1-5.8S rDNA-ITS2)

ITS2 영역의 유전적 유연관계분석
JNCPCRI 병귤 품종과 GenBank에 등록된 재래감귤 11품종 의 ITS2 영역에 대한 염기서열 분기값(Sequence divergency)을 계산한 결과, 빈귤(JQ990189), 당유자(JQ990179), 홍귤 (JQ990178), 편귤(JQ990181) 품종은 0.00886로 다른 재래감 귤 품종들에 비하여 염기서열의 다양성 정도가 가장 낮았다 (Table
4). 즉 JNCPCRI 병귤 품종의 염기서열은 이 4품종의 염기서열과 비슷하며 유전적으로 유사하다는 것이 확인되었다 (Table
4). 반면, GenBank에 등록된 병귤(JQ990189) 품종은 염기서열의 분기값이 0.04073으로 재래감귤 11품종 중 염기서 열 다양성 정도가 3번째로 높았다(Table
4).
Yao et al. (2010)의 보고에 따르면 ITS2 영역은 염기서열이 변화가 높게 나타나기 때문에 식물의 계통분류와 발생연구에 있어서 ITS1보다는 ITS2 영역의 염기를 이용한 연구들이 수행되고 있는데(
Raveendar et al. 2015a,
Raveendar et al. 2015b), 본 연구 결과에서도 ITS1 영역 보다는 ITS2 영역의 유전적인 변화가 많다는 것을 알 수 있었다. 따라서, 감귤의 계통분류와 발생연구에 있어서 ITS2 영역을 이용한 연구가 바람 직 하다고 생각된다. 그리고, ITS2 영역에 대한 염기들을 이용하여 JNCPCRI 병귤 품종과 재래감귤 품종간의 유연관계를 분석한 결과 Group I과 Group II의 2개의 분류군으로 나눌 수 있었고, Group I는 다시 3개의 분계조 나눌 수 있었다. JNCPCRI 병귤 품종은 Ia에 속하여 있으며, 빈귤(JQ990180), 당유자(JQ990179), 홍귤 (JQ990178), 편귤(JQ990181) 품종과 유전적으로는 가장 근연 관계에 있음이 확인되었다. 하지만, GenBank에 등록된 병귤 (JQ990189) 품종은 동정귤(JQ990187)과 함께 Ic에 속하며 JNCPCRI 병귤 품종과는 유전적으로 유연관계가 멀다는 것이 확인되었다(Fig.
3B). 모든 재래감귤은 화분수정에 의하여 많은 종자를 생산할 수 있는 자가합성 품종들(
Chae et al. 2012,
Song et al. 2011,
Yun et al. 2007)이며, 화분수정으로 인하여 유전적 으로 다른 실생묘를 생산할 수 있는 유성배를 갖고 있다(
Dhillon et al. 1993). 또한, 이들 품종들은 주변에 재배되고 있는 화분 임성의 높은 품종들과의 수정을 통하여 종자를 형성할 수 있는 자가합성 품종들이다(
Chae et al. 2012,
Song et al. 2011). 따라서 본 결과처럼 JNCPCRI 병귤 품종의 ITS2 영역 유전자가 빈귤(JQ990180), 당유자(JQ990179), 홍귤(JQ990178), 편귤 (JQ990181) 품종들과 유전적으로는 근연관계에 있다는 것은 이들 품종들과 수정을 통하여 변이가 생긴 것으로 판단 할 수 있다.
Table 4.Sequence divergences in the ITS2 gene regions of JNCPCRI cultivar and related Jeju Native Citrus Varieties.
Table 4.
|
No. |
Cultivars |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
|
1 |
C. aurantium (JQ990184) |
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
C. benikoji (JQ990186) |
0.08955 |
|
|
|
|
|
|
|
|
|
|
|
|
3 |
C. erythrosa (JQ990187) |
0.04540 |
0.10896 |
|
|
|
|
|
|
|
|
|
|
|
4 |
C. leiocarpa (JQ990180) |
0.03160 |
0.09425 |
0.03614 |
|
|
|
|
|
|
|
|
|
|
5 |
C. grandis (JQ990179) |
0.03160 |
0.09425 |
0.03614 |
0.00000 |
|
|
|
|
|
|
|
|
|
6 |
C. nippokoreana (JQ990183) |
0.04088 |
0.10438 |
0.04080 |
0.00886 |
0.00886 |
|
|
|
|
|
|
|
|
7 |
C. platymamma (JQ990189) |
0.03610 |
0.09890 |
0.02697 |
0.03149 |
0.03149 |
0.03610 |
|
|
|
|
|
|
|
8 |
C. pseudogulgul (JQ990185) |
0.08468 |
0.01335 |
0.10391 |
0.08933 |
0.08933 |
0.09940 |
0.09393 |
|
|
|
|
|
|
9 |
C. sunki (JQ990188) |
0.02697 |
0.08915 |
0.04549 |
0.03150 |
0.03150 |
0.04075 |
0.03614 |
0.08426 |
|
|
|
|
|
10 |
C. tachibana (JQ990178) |
0.03160 |
0.09425 |
0.03614 |
0.00000 |
0.00000 |
0.00886 |
0.03149 |
0.08933 |
0.03150 |
|
|
|
|
11 |
C. tangerina (JQ990181) |
0.03160 |
0.09425 |
0.03614 |
0.00000 |
0.00000 |
0.00886 |
0.03149 |
0.08933 |
0.03150 |
0.00000 |
|
|
|
12 |
JNCPCRI |
0.04088 |
0.10438 |
0.04543 |
0.00886 |
0.00886 |
0.01784 |
0.04073 |
0.09940 |
0.04075 |
0.00886 |
0.00886 |
|
전체ITS 영역의 유전적 유연관계분석
JNCPCRI 병귤 품종과 GenBank에 등록된 재래감귤 11품종 의 전체 ITS 유전자 영역에 대한 염기서열 분기값(Sequence divergency)을 계산한 결과, 홍귤(JQ990178) 품종이 JNCPCRI 병귤 품종과의 염기서열 분기값이 0.00483로 다른 품종들에 비하여 염기서열 다양성 정도가 가장 낮았으며, 그 다음으로 빈귤(JQ990180) 그리고 편귤(JQ990181) 2품종이 0.00644, 당 유자(JQ990179) 품종이 0.00806 그리고 청귤(JQ990183) 품종 이 0.00968 순으로 낮았다(Table
5). 반면에 병귤(JQ990189) 품종이 0.01456으로 JNCPCRI 병귤 품종과의 염기서열 다양성 정도가 재래감귤 11품종 중 6번째로 높게 나타났다(Table
5). 결과적으로 이들 재래감귤 11품종 중 홍귤(JQ990178) 품종은 다른 재래감귤 품종들에 비하여 JNCPCRI 병귤 품종간에 염기 구성이 가장 비슷한 것으로 나타났다(Table
5), 위 결과는 ITS2 영역의 염기를 이용한 유연관계 결과와 유사하였다.
Table 5.Sequence divergences in the total ITS gene regions of JNCPCRI cultivar and related Jeju Native Citrus Varieties.
Table 5.
|
No. |
Cultivars |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
|
1 |
C. aurantium (JQ990184) |
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
C. benikoji (JQ990186) |
0.10613 |
|
|
|
|
|
|
|
|
|
|
|
|
3 |
C. erythrosa (JQ990187) |
0.01948 |
0.10925 |
|
|
|
|
|
|
|
|
|
|
|
4 |
C. leiocarpa (JQ990180) |
0.01788 |
0.11158 |
0.01948 |
|
|
|
|
|
|
|
|
|
|
5 |
C. grandis (JQ990179) |
0.01951 |
0.11341 |
0.02113 |
0.00160 |
|
|
|
|
|
|
|
|
|
6 |
C. nippokoreana (JQ990183) |
0.02117 |
0.11536 |
0.02113 |
0.00321 |
0.00482 |
|
|
|
|
|
|
|
|
7 |
C. platymamma (JQ990189) |
0.01620 |
0.10944 |
0.01293 |
0.01456 |
0.01620 |
0.01620 |
|
|
|
|
|
|
|
8 |
C. pseudogulgul (JQ990185) |
0.09859 |
0.02776 |
0.10173 |
0.10399 |
0.10580 |
0.10772 |
0.10189 |
|
|
|
|
|
|
9 |
C. sunki (JQ990188) |
0.01955 |
0.10591 |
0.02278 |
0.02448 |
0.02613 |
0.02781 |
0.02279 |
0.09838 |
|
|
|
|
|
10 |
C. tachibana (JQ990178) |
0.01624 |
0.10976 |
0.01784 |
0.00160 |
0.00321 |
0.00482 |
0.01293 |
0.10219 |
0.02284 |
|
|
|
|
11 |
C. tangerina (JQ990181) |
0.01788 |
0.11158 |
0.01948 |
0.00000 |
0.00160 |
0.00321 |
0.01456 |
0.10399 |
0.02448 |
0.00160 |
|
|
|
12 |
JNCPCRI |
0.01788 |
0.11158 |
0.01948 |
0.00644 |
0.00806 |
0.00968 |
0.01456 |
0.10399 |
0.02448 |
0.00483 |
0.00644 |
|
이전 연구자들의 보고에 따르면 병귤 품종은 당유자 (JQ990179), 편귤(JQ990181), 홍귤(JQ990178), 빈귤 (JQ990180) 품종들은 사두감(JQ990185), 빈귤(JQ990180) 품 종들과 염기서열 구성이 가장 유사하다고 하였는데(
Jung et al. 2005,
Sun et al. 2015), 본 연구에서도 JNCPCRI 병귤 품종은 홍귤(JQ990178), 빈귤(JQ990180), 편귤(JQ990181), 당유자 (JQ990179) 그리고 청귤(JQ990183) 품종과 유전적으로 가깝 다는 점에서 이전 연구자들(
Jung et al. 2005,
Sun et al. 2015)의 결과와 비슷하였다. 하지만 본 연구 결과에서 얻어진 JNCPCRI 병귤 품종은 이전 연구자(
Sun et al. 2015)가 GenBank에 등록한 병귤(JQ990189) 품종과는 염기서열 분기값이 0.01456로 염기 서열 다양성 정도가 GenBank에 등록된 재래감귤 11품종 중 6번째로 높은 결과를 얻었다(Table
5). 이와 같은 결과는 ITS2 영역의 유전적인 차이로 인하여 형태학적으로 동일한 품종이지 만 유전적으로 차이를 보인 것이라 판단 할 수 있었다.
전체 ITS 유전자 영역에 대한 염기들을 이용하여 JNCPCRI 병귤 품종간의 근연관계를 조사한 결과 Group I과 Group II의 2개 분류군과 4개의 분계조로 나눌 수 있었다. Group I에는 빈귤(JQ990180), 편귤(JQ990181), 당유자(JQ990179), 홍귤 (JQ990178), 청귤(JQ990183), JNCPCRI 병귤, 동정귤 (JQ990187), 병귤(JQ990189), 지각(JQ990184) 그리고 진귤 (JQ990188) 품종으로 구성되어 있으며, Group II에는 감자 (JQ990186) 그리고 사두감(JQ990185) 품종의 그룹으로 구성 되어 있다(Fig.
3C).
또한, Group I은 4개의 분계조로 다시 나눌 수 있는데, 빈귤 (JQ990180), 편귤(JQ990181), 당유자(JQ990179), 홍귤 (JQ990178), 청귤(JQ990183), JNCPCRI 병귤 품종들은 Ia, 동정귤(JQ990187)과 병귤(JQ990189) 품종은 Ib, 지각 (JQ990184) 품종은 Ic 그리고 진귤(JQ990188) 품종은 Id로 나눌 수 있었다. 비록, JNCPCRI 병귤과 GenBank 등록된 병귤 (JQ990189) 품종은 동일한 GroupI에 속하나 다른 분계조에 속하여 있는 점으로 보아 유전적으로는 근연관계가 멀다는 것이 확인 되었다. 반면에, Ia에 속하는 빈귤(JQ990180), 편귤 (JQ990181), 당유자(JQ990179), 홍귤(JQ990178) 그리고 청귤 (JQ990183) 품종은 JNCPCRI 병귤 품종과는 형태학적으로는 서로 다르나 유전적으로 근연관계에 있으며, 이 중 청귤 (JQ990183) 품종은 유전적으로 가장 근연관계에 있음이 확인되 었다(Fig.
3C). 또한, JNCPCRI 병귤 품종은 빈귤(JQ990180) 그리고 편귤(JQ990181) 품종들로부터 기원된 것으로 생각되며, Ia에 속하는 품종들간의 오랜 기간 동안 재배되는 과정에서 수정 을 통하여 유전적으로 변이가 발생한 것으로 생각된다(
Dhillon et al. 1993,
Sun et al. 2015). 모계 유전과 관련된 엽록체 trnL-trnF 스페이서 영역을 이용하여 GenBank에 등록된 재래감 귤 11품종과 유연관계를 비교 분석하였을 때 병귤 품종은 독립된 단일 품종으로 구성되어 있다고 하였다(
Jung et al. 2005). 하지 만, 본 연구에서는 JNCPCRI 병귤 품종이 GenBank에 등록된 유자(AY321663), 감자(AY321666), 진귤(Y321665) 품종들 과 근연관계가 가까웠고, 지각(AY321661) 품종과는 근연관계 가 가장 멀었다. 또한, 병귤(AY321659) 품종과는 염기의 변이를 보이는 site가 372개 중 4개 나타났으며, 근연관계 또한 위 3 품종(유자, 감자, 진귤)들에 비하여 멀게 나타났다(자료 미제시). 위 결과로 병귤 품종은 모계 유전 된 것이 아니라 수정을 통한 종자 번식된 것으로 판단 되었다. 따라서, JNCPCRI 병귤 품종은 GenBank에 등록된 병귤(JQ990189) 품종과 유전적으로 유연관 계가 멀어진 것이 아닌가 판단할 수 있었다.
ITS 영역을 이용하여 감귤에 대한 유연관계를 분석한
Sun et al. (2015)등에 의하면 당유자(JQ990179) 품종을 제외한 나머 진 재래감귤 품종들은
Sinocitrus종에 속하며, 재배지역에 관계 없이 재래감귤들은 유전적으로 근연관계가 비슷하다고 하였다. 그러나 GenBank에 등록된 병귤(JQ990189) 품종과 본 연구소 내에서 재배되고 있는 병귤(JNCPCRI) 품종간에는 유전적으로 근연관계가 Ia 분계조에 속하는 품종들에 비하여 멀다는 점에서 같은 모계에서 분리 증식된 것이 아니라 종자번식을 통하여 오랜 기간 동안 번식되어 왔기 때문에 유전적으로 변이가 생긴 것으로 생각된다. 따라서, 차후 제주도내에 보존 되고 있는 병귤 품종들에 대한 정확한 기원 규명을 위하여 유전적인 근연관계를 조사 할 필요가 있다고 판단된다.
적 요
제주도내 재래감귤 12품종들에 대한 기원은 대부분 중국으로 부터 유래 한 것으로 사료되고 있다. 그러나, 병귤(Citrus platymamma Hort. ex Tanaka) 품종에 대한 기원은 정확하게 알려진 바가 없다. 따라서, 본 연구는 재래감귤 병귤 품종에 대한 기원 규명 과 재래감귤 품종들 간의 계통분류학적 연구 목적으로, 최근 식물 계통분류학적 유연관계 분석에 이용되고 있는 핵 리보솜 DNA내 ITS 영역의 프라이머를 디자인하여 PCR를 수행하였다. JNCPCRI(Jeju Native Citrus platymamma Citrus Research Institute) 병귤의 ITS1 유전자 영역은 247bp 였고 ITS2 유전자 영역은 228bp, 전체 ITS 유전자 영역 (ITS1-5.8S-ITS2)은 638bp 였다. JNCPCRI 병귤의 ITS1 유전 자 영역 염기서열 분석을 기초로 하여 유연관계를 분석한 결과 JNCPCRI 품종은 GenBank에 등록된 병귤(JQ990189) 품종과 유전적으로 동일하였고, ITS2 유전자 영역은 빈귤(JQ990180), 홍귤(JQ990178), 당유자(JQ990179) 그리고 편귤(JQ990181) 품종과 유전적으로 비슷하였으며, 전체 ITS 유전자 영역은 홍귤 (JQ990178) 품종과 유전적으로 비슷하였다. 또한, 전체 ITS 유전자 영역으로 JNCPCRI 품종의 계통학적 근연관계를 분석한 결과 청귤(JQ990183) 품종과 가장 근연관계에 있으며, 빈귤 (JQ990180) 그리고 편귤(JQ990181) 품종으로부터 기원된 것 으로 조사되었다. 반면에, JNCPCRI 품종은 병귤(JQ990189) 품종과 형태학적으로 동일하나 ITS 영역은 유전적으로 상이성 을 보였다. 위 결과로 JNCPCRI 품종은 주변 품종들간의 수정을 통하여 ITS 영역이 유전적으로 변이가 생긴 것으로 사료된다.
사 사
본 연구는 농촌진흥청 PJ01027304 사업과 박사후 연구원 지원 프로그램에 의해 수행되었음.
References
- 1. Baldwin BG, Sanderson MJ, Porter JM, Wojciechowski MF, Campbell CS, Donoghue MJ. The ITS region of nuclear ribosomal DNA A valuable source of evidence on angiosperm phylogeny. Ann. Mo. Bot. Gard 1995. 82: 247-277.
- 2. Barrett HC, Rhodes AM. A numerical taxonomic study of affinity relationships in cultivated Citrus and its close relatives. Syst. Bot 1976. 1: 105-136.
- 3. Chae CW, Yun SH, Park JH, Kim MJ, Koh SW, Song KJ, Lee DH. Frequency of spontaneous polyploids inmonoembryonic Jeju native citrus species and some mandarin cultivars. Journal of Life Science 2012. 22: 871-879.
- 4. Cheng Y, Vicente MC, Meng H, Guo W, Tao N, Deng X. A set of primers for analyzing chloroplast DNA diversity in Citrus and related genera. Tree Physiol 2005. 25: 661-672.
- 5. Deng ZN, Gentile A, Nicolosi E, Domina F, Vardi A, Tribulato E. Identification of in vivo and in vitro lemon mutants by RAPD marker. J. Hortic. Sci 1995. 70: 117-125.
- 6. Deng ZN, Malfa S, La Xie XM, Xiong XG, Gentile A. Identification and evaluation of chloroplast uni-and trinucleotide sequence repeats in citrus. Sci. Hortic 2007. 111: 186-192.
- 7. Dhillon RS, Kaundal GS, Cheema SS. Nucellar embryony for propagating Citrus. Indian Hortic 1993. 38: 44-45.
- 8. Federici CT, Fang DQ, Scora RW, Roose ML. Phylogenetic relationships within the genus Citrus (Rutaceae) and related genera as revealed by RFLP and RAPD analysis. Theor. Appl. Genet 1998. 96: 812-822.
- 9. Froelicher Y, Mouhaya W, Bassene JB, Costantino G, Kamiri M, Luro F, Morillon R, Ollitrault P. New universal mitochondrial PCR markers reveal new information on maternal citrus phylogeny. Tree Genetics & Genomes 2011. 7: 49-61.
- 10. Gulsen O, Roose ML. Lemons diversity and relationships with selected Citrus cultivars and species and comparison with related genera. Theor. Appl. Genet 2001. 72: 170-177.
- 11. Hall TA. Bio Edit a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT Nucl. Acids Symp. Ser 1999. 41: 95-98.
- 12. Hazarika TK, Hazarika BN, Shukla AC. Genetic variability and phylogenetic relationships studies of genus Citrus L with the application of molecular markers. Genet. Resour Crop Evol 2014. 61: 1441-1454.
- 13. Herrero R, Asins MJ, Pina JA, Carbonell EA, Navarro L. Genetic diversity in the orange subfamily Aurantioideae II Genetic relationships among genera and species. Theor. Appl. Genet 1996. 93: 1327-1334.
- 14. Hirai M, Kozaki I, Kajiura I. Isozyme analysis and phylogenic relationship of Citrus. Japan. J. Breed 1986. 36: 377-389.
- 15. Hynniewta M, Malik SK, Rao SR. Genetic diversity and phylogenetic analysis of Citrus (L) from north-east India as revealed by meiosis, and molecular analysis of internal transcribed spacer region rDNA. Meta Gene 2014. 237-251.
- 16. Hyun JM, Park KJ, Kim SS, Park SM, Lee YJ, An HJ. Antioxidant and Anti-inflammatory Effects Solvent Fractions from the Peel of the Native Jeju Citrus ‘’Hongkyool’ and ‘Pyunkyoo’. Journal of Life Science 2015. 25: 1132-1138.
- 17. Jung YH, Kwon HM, Kang SH, Kang JH, Kim SC. Investigation of the phylogenetic relationships within the genus Citrus (Rutaceae) and related species in Korea using plastid trnL-trnF sequences. Sci. Hortic 2005. 104: 179-188.
- 18. Kim CM, Kim KS, Kim MH, Huh IO. Taxonomical and Phytochemical Studies of Citrus Plants Native to Je Ju Island (I) Flavour Patterns of the Citrus Peel Oils and One of the Citrus Flavonoids. J. Pharmacogn 1979. 10: 13-16.
- 19. Kim YC, Koh KS, Koh JS. Changes of flavonoids in the peel of Jeju native citrus fruits during maturation. Food Sci. Biotechnol 2001. 10: 483-487.
- 20. Kim YD, Ko WJ, Koh KS, Jin JY, Hyun KS. Composition of flavonoids and antioxidative activity from juice of Jeju native citrus fruits during maturation. Korean. J. Nutr 2009. 42: 278-290.
- 21. Kim YD, Mahinda S, Koh KS, Jeon YJ, Kim SH. Reactive oxygen species scavenging activity of Jeju native citrus peel during maturation. J. Korean Soc. Food Sci. Nutr 2009. 38: 462-469.
- 22. Kim HY. Distribution, taxonomy, horticultural characters of the local Citrus spp. in Cheju, and the genetic markers among them. Ph.Diss., Cheonnam National University., Korean 1988.
- 23. Ko WC, Lee SR. Effect of immature Citrus sunki peel extract on neuronal cell death. Korean. J. Med. Crop Sci 2015. 23: 144-149.
- 24. Luro F, Costantino G, Terol J, Argout X, Allario T, Wincker P, Talon M, Ollitrault P, Morillon R. Transferability of the EST-SSRs developed on Nules Clementine (Citrus clementina Hort ex Tan) to other Citrus species and their effectiveness for genetic mapping. BMC Genomics 2008. 9: 1471-2164.
- 25. Moon DK, Ko KC. Isozymes as genetic markers in Citrus growing in Cheju and their use for dentification of nucellar and zgotic seedlings II PGI, PGM, IDH, GOT and MDH isozymes. J. Korean Soc. Hortic. Sci 1992. 33: 132-145.
- 26. Oh JB. Classification of Citrus growing in Cheju by RAPD and selection of markers for species or cultivars identification. M.S. Thesis Cheju National University Korean 1996.
- 27. Olmstead RG, Palmer JD. Chloroplast DNA systematic a review of methods and data analysis. Am. J. Bot 1994. 81: 1025-1224.
- 28. Penjor T, Yamamoto M Uehara M, Ide M, Matsumoto N, Matsumoto R, Nagano Y. Phylogenetic relationships of Citrus and its relatives based on matK gene sequences. PloS one 2013. 8: e62574.
- 29. Pessina D, Gentil R
, Barcaccia G, Nicolè S, Rossi G, Barbesti S, Sgorbati S. DNA content, morphometric and molecular marker analyses of Citrus limonimedica C limon and C medica for the determination of their variability and genetic relationships within the genus Citrus. Sci. Hortic 2011. 129: 663-673.
- 30. Rahman MM, Nito N, Isshiki S. Cultivar identification of ‘Yuzu’ (Citrus junos Sieb Ex Tanaka) and related acid Citrus by leaf isozymes. Sci. Hortic 2001. 87: 191-198.
- 31. Raveendar S, Lee Jr, Park JW, Lee GA, Jeon YA, Lee YJ, Cho GT, Ma KH, Lee SY, Chung JW. Potential use of ITS2 and matK as a two-locus DNA barcode for identification of Vicia species. Plant Breed. Biotech 2015. 3: 58-66.
- 32. Raveendar S, Lee Jr, Park JW, Lee GA, Jeon YA, Lee WH, Cho GT, Ma KH, Lee SY, Chung JW. Identification of genus Vigna using ITS2 and matK as a two-locus DNA barcode. Plant Breed. Biotech 2015. 3: 153-159.
- 33. Scora RW. On the history and orgin of citrus. Bull. Torrey Bot. Club 1975. 102: 369-375.
- 34. Song KJ, Kim SB, Park JH, Oh EU, Lee KG, Kim DW, Kang JH, Kim JS, Oh JH, Gmitter FG. Frequencyand growth characteristics of polyploids occurred spontaneously in some mandarin hybrids. Kor. J. Hort.Sci. Technol 2011. 29: 617-622.
- 35. Sun YL, Kang HM, Han SH, Park YC, Hong SK. Taxonomy and phylogeny of the genus citrus based on the nuclear ribosomal dna its region sequence. Pak. J. Bot 2015. 47: 95-101.
- 36. Tingey SV, Tufo JP. Genetic analysis with random amplified polymorphic DNA markers. Plant Physiol 1993. 101: 349-352.
- 37. USDARS (United States Department of Agricultural Rese arch Service)U.S. National Plant Germplasm Sy stem. http://www.ars-grin.gov/npgs/index.html# (accessed March 29 2016 ) 2016.
- 38. Xu CJ, Bao L, Zhang B, Bei ZM, Ye XY, Zhang SL, Chen KS. Parentage analysis of huyou (Citrus changshanensis) based on internal transcribed spacer sequences. Plant Breed 2006. 125: 519-522.
- 39. Yao H, Song JY, Liu C, Luo K, Han JP, Li Y, Pang XH, Xu HX, Zhu YJ, Xiao PG, Chen SL. Use of ITS2 Region as the Universal DNA Barcode for Plants and Animals. PloS one 2010. 5: e13102.
- 40. Yang EJ, Lee HJ, Kang GJ, Park SS, Yoon WJ, Kang HK, Cho SK, Yoo ES. Anti-inflammatory Effect of Dangyuja (Citrus grandis Osbeck) Leaves in LPS-stimulated RAW 264. Cells. Food Sci. Biotechnol 2009. 18: 1063-1070.
- 41. Yu EA, Kim HG, Song Y, Kim GS, Kim JH, Kim DY, Jin JS, Lee SJ, Sung NJ, Shin SC. Determination of flavonoids in the peel of Jingyul (Citrus sunki Hort Ex Tanaka) using a HPLC-MS/MS and the antioxidant activity. J. Agric. Life Sci 2014. 48: 227-234.
- 42. Yun JU, Yang HB, Jung YH, Yun SH, Kim KS, Kim CS, Song KJ. Identification of Zygotic and Nucellar Mandarin Seedlings Using Randomly Amplified Polymorphic DNA. Hort. Environ. Biotechnol 2007. 48: 171-175.
- 43. Whitkus R, Doeet J, Wendel F. In: Philps RL, Vasil JK. (Eds). Nuclear DNA markers in systmatics and evolution DNA-based markers in plants. 1994. The Netherlands: Kluwer Academic Publ Dordrecht, pp. 116-141.