적요
A comprehensive evaluation of 515 Korean wheat germplasms, including cultivars, experimental lines, and landraces, was conducted over 2 years under upland field conditions to characterize major agronomic and grain traits. Allelic variation at 13 key functional loci was assessed using Kompetitive Allele-Specific PCR (KASP) and PCR-based markers. The winter-type vrn-A1 with a single copy (CNV=1; 40.2%) advanced heading by approximately 5 days compared to multiple-copy genotypes, and winter-type vrn-B1 (88.5%) advanced heading by 2 days compared to the spring-type. The photoperiod-insensitive alleles Ppd-B1a (5.6%) and Ppd-D1a (76.3%) advanced heading by 3 and 4 days, respectively, with a combined effect of up to 6 days. Semi-dwarfing alleles Rht-B1b and Rht-D1b showed reduced culm lengths of 2.1 cm and 4.7 cm, respectively, and the Rht-B1a/Rht-D1b genotype was 6 cm shorter than Rht-B1a/Rht-D1a. The Pina-D1a/Pinb-D1a genotype had the lowest kernel hardness value (32.2), whereas Pina-D1b/Pinb-D1a had the highest (60.5). The thousand kernel weight ranged from 36.1 mg to 42.5 mg depending on the allelic combinations of TaCwi-A1, TaGW2-6A, and TaSus2-2B. Cultivars and experimental lines were clearly distinguished from landraces based on phenotype-based clustering, with the majority of cultivars (81.6%) and experimental lines (68.3%) grouped into cluster III. In contrast, landraces were predominantly distributed in clusters I (55.1%) and II (29.2%). Random forest analysis identified four genes, Ppd-D1, Pina-D1, Pinb-D1, and WAPO-A1, as major contributors to cluster classification. Cluster III was highly enriched with alleles favorable for earliness (Ppd-D1a, 98.3%) and grain hardness (Pina-D1b and Pinb-D1b, 57.9%). WAPO-A1b, an allele associated with an increased spikelet number per spike, was more frequently observed in clusters I (94.6%) and II (79.1%) than in cluster III (58.4%).
서언
국산 밀 자급률 제고를 위해 제정된 “밀산업육성법”은 국산 밀의 가격 경쟁력 강화, 생산 기반 확충, 품질 고급화, 유통 차별화와 안정적 소비 시장 확보 등을 포함하고 있다(
Jung et al. 2020). 국산 밀 자급률 달성을 위해 생산 및 유통 기반과 정책 지원을 통한 가격 경쟁력 확보도 중요하지만, 소비 활성화를 위한 품질 고급화가 필수적이다. 국산 밀의 수량성 증진은 농업인 및 생산 단체의 안정적 소득 보장을 위해 필요하며, 소비자가 만족하며 수입산과 경쟁할 수 있는 품종 개발을 위해서는 농업적 형질과 품질 특성이 개선된 품종의 지속적 개발이 필요하다.
밀 육종 프로그램에서는 수량성과 품질 개선을 위해 다양한 유전자원이 활용된다. 외국 유전자원은 유용한 자원을 도입하고 다양성을 확보하는데 유리한 점이 있지만, 국내 재배 환경 적응력이 떨어질 가능성이 있어, 품종화를 위해서는 추가적인 적응 기간이 필요하여 상대적으로 육종 과정이 길어질 수 있다. 반면, 재래종을 포함한 국내 유전자원은 국내 기후, 토양, 병해충 등에 적응되어 있어, 외국 자원에 비해 유전적 다양성은 다소 제한적일 수 있지만, 국내 재배 환경에 적합한 고품질 및 고수량 품종 육성에 적합하다(
Son et al. 2021).
유전자원을 효과적으로 활용하고 육종 효율성과 정확성을 높이기 위해 분자표지를 활용한 MAS (Marker-Assisted Selection)가 밀 육종에서 널리 활용되고 있다(
Gupta et al. 2010,
Liu et al. 2012,
Maccaferri et al. 2022). 출수기는 저온 요구도와 광주기 반응성에 따라 조절되며, Vernalization 유전자군(
Vrn-1,
Vrn-2,
Vrn-3)과 Photoperiod 유전자군(
Ppd-1)이 주요한 유전적 요인으로 알려져 있다(
Kamran et al. 2014). 특히
Vrn-A1에서 겨울형 대립유전자의 복제수가 증가하면 더 긴 춘화처리가 필요하며, 출수가 지연된다(
Díaz et al. 2012). 우리나라에서 밀은 대부분 벼나 콩과 이모작 재배를 주로 하기 때문에 생육시기 및 농작업 경합을 피하기 위해 출수가 빠른 조기 수확 품종 육성이 중요하다. 간장은 도복과 연관이 있으며, 녹색혁명 반왜성 표현형과 관련된
Rht-B1과
Rht-D1 돌연변이형이 널리 보고되어 있다(Pearce 2011). 일수립수는 수량구성요소 중 유전력이 높은 표현형으로
WAPO-A1 유전자가 7AL QTL의 원인 유전자로 밝혀지며, 이삭 구조 및 일수립수 조절에 중요한 역할을 하는 것으로 알려져 있다(
Kuzay et al. 2022). 종실경도는 제분 후 밀가루의 손상전분과 가공시 수분흡수율에 영향을 미치는데 5D 염색체의
Pina-D1과
Pinb-D1 변이에 의해 조절되며, 대립유전자 조합에 따라 연질⋅경질밀로 구분된다(
Morris 2002). 종실 크기 및 무게는 수량과 제분율에 큰 영향을 미치는며
TaCwi-A1,
TaGW2-6A,
TaSus2-2B와 같은 유전자들이 조절하는데 다양한 집단에서 천립중 및 수량성 QTL로 반복적으로 보고되고 있다(
Gao et al. 2023).
국산 밀에 대해서도 주요 유전자에 대한 분석이 일부 수행되었으나(
Cho et al. 2015a,
Cho et al. 2015b,
Kim et al. 2020), 대부분 품종에 국한되어 있어, 국내 재래종과 육성 계통을 포함한 전반적인 유전적 다양성 및 선발 흐름을 파악하기에는 한계가 있었다. 따라서 본 연구는 국산 밀 유전자원 515점(품종 49점, 육성 계통 123점, 재래종 343점)을 대상으로 출수기, 간장, 수장, 종실경도, 천립중 등의 주요 형질과 관련된 13개 유전자의 대립유전자 변이를 분석하고, 형질과의 연관성을 평가하였다. 또한 표현형 기반 군집분석 및 랜덤포레스트 기법을 통해 군집 간 차이를 설명하는 핵심 유전자를 선발함으로써, 국내 밀 육종에 실용적인 기초자료를 제공하고자 하였다.
재료 및 방법
공시재료
본 연구에는 농촌진흥청 국립식량과학원 밀육종연구실에서 보유하고 있는 국내 밀 유전자원515점을 이용하였다. 유전자원515점은 품종 49점, 계통 123점과 재래종 343점을 포함하고 있으며, 재래종에는 북한자원 77점이 포함되었다. 공시재료는 전작 조건으로 2년간(2022년과 2023년) 국립식량과학원 전작 포장에서 농촌진흥청 표준재배법(
RDA 2012)에 준하여 재배하였다. 공시재료는 완전 임의 배치법으로 시험포장에서 3 반복으로 0.25 m×4 m×3열 조건에서 재배하였으며, 10월 25일에 파종하여 다음 해 6월 20일에 수확하였다. 2022년과 2023년 평균 기온은 각각 9.3℃와 9.8℃로 평년 평균 기온(8.8℃)보다 높았으며, 강수량은 2022년에는 342 mm로 평년 강수량(457 mm)보다 낮았지만, 2023년은 458 mm로 평년과 차이가 없었다.
농업형질 조사는 농촌진흥청 농업 과학 기술 조사 분석 기준(
RDA 2012)에 준하여 실시하였다. 출수기는 전체 단위 면적당 이삭 수 중에 40%가 출수한 날로 하였으며, 간장은 지면에서 이삭목까지 길이로 나타내었고, 수장은 이삭목에서 이삭 끝까지 길이를 측정하였다. 원맥 특성은 수확 후 한달 간 비닐하우스에서 수분 함량이 12% 이하가 되도록 건조하였고, 탈곡 후 2.0 mm 체로 정선된 건전립을 10℃ 저온실에 보관하였다. Single Kernel Characterization System (SKCS Model 4100, Perten Instruments, Springfield, IL, USA)을 이용하여 종실 300립의 경도와 무게를 측정하고, 이를 평균값으로 나타내었다(
Bettge & Morris 2000).
온실에서 3주간 생육시켜 자란 식물체의 3엽을 채취하여 액체 질소로 급속 냉동시킨 후 유발을 이용하여 미세분말로 분쇄한 뒤 -70℃에 보관하였다. Genomic DNA는 잎 분말(100 mg)로부터 식물용 genomic DNA 분리 시약(Solgent, Korea)을 이용하여 추출하였다. 추출한 genomic DNA는 Nanodrop 1000 spectrophotometer (Thermo Scientific, Wilmington, USA)를 사용하여 정량하였고, -20℃에 보관하여 사용하였다.
유전자형 분석은 파성, 광주기 반응, 간장, 일수립수, 종실크기, 종실경도와 관련된 13개 유전자를 대상으로 수행하였다. 이 중
Vrn-A1,
WAPO-A1와
Pinb-D1 유전자는 KASP (Kompetitive Allele Specific PCR) 마커를 활용하여 분석하였으며(Supplementary Table 1), 나머지 유전자는 PCR 기반 마커 분석을 실시하였다. 파성 유전자인
Vrn-A1과
Vrn-B1의 대립유전자 조성은 각각
Yan et al. (2004) 및
Fu et al. (2005)의 방법을 따랐으며,
Vrn-A1의 copy number variation 분석을 위해 KASP 마커를 추가로 적용하였다(
Diaz et al. 2012). 광주기 반응 관련 유전자
Ppd-A1,
Ppd-B1과
Ppd-D1은
Bentley et al. (2011)과
Seki et al. (2011)의 방법에 따라 분석하였다. 간장 관련 유전자인
Rht-B1과
Rht-D1은
Ellis et al. (2002) 및
Zhang et al. (2006)의 방법에 따라 분석하였고, 일수립수에 관여하는
WAPO-A1 유전자는
Kuzay et al. (2022) 및
Zhou et al. (2023)의 방법을 참고하여 PCR 기반 마커와 KASP 마커를 병행해 분석하였다. 종실 특성과 관련된
TaCwi-A1,
TaGW2-6A와
TaSus2-2B는 각각
Ma et al. (2012),
Su et al. (2011) 및
Jiang et al. (2010)의 방법에 따라 분석하였다.
Pina-D1 유전자는
Gautier et al. (1994)을 참고하여 PCR로 분석하였고,
Pinb-D1 유전자는 Polymarker를 통해 제작한 KASP 마커를 활용하여 분석하였다.
KASP 분석을 위해 SNP가 3' 말단에 위치하도록 표준 FAM 형광서열(5'-GAAGGTGACCAAGTTCATGCT-3') 및 HEX 형광서열(5'-GAAGGTCGGAGTCAACGGATT-3')을 포함한 대립유전자 특이적 프라이머를 설계하였으며, 96-well 형식의 반응 조건은 총 10 µL (DNA 5 µL, 2× KASP Master Mix 5 µL, Assay mix 0.14 µL)로 구성하였다. 반응은 CFX96 Real-Time Thermal Cycler (Bio-Rad Laboratories, USA)를 사용하여 제조사에서 권장하는 터치다운 PCR 조건으로 수행하였고, CFX Maestro Software 2.0을 이용해 유전자형을 판독하였다. PCR 기반 마커 분석은 Veriti® Dx 96-Well Thermal Cycler (Applied Biosystems, USA)를 사용하였으며, 생성된 PCR 산물은 1.5% agarose gel에서 135 V 조건으로 1시간 전기영동 후, UV transilluminator를 통해 확인하였다.
통계분석
통계분석은 R (Version 4.3.1)을 이용하여 수행하였다. 기술통계, 분산분석(ANOVA, Analysis of Variance), Duncan's 다중검정(DMRT, Duncan's Multiple Range Test)은 agricolae 패키지로 분석하였으며, 유전형은 임의효과, 재배연도는 고정 효과로 설정하여 lme4 패키지를 통해 형질별 BLUPs (Best Linear Unbiased Predictions)를 산출하고 대표 표현형으로 활용하였다. 표현형 분포는 ggplot2와 dplyr 패키지로 시각화하였고, 형질간 상관관계는 Hmisc 패키지의 rcorr () 함수를 이용해 피어슨 상관계수와 유의확률을 계산하였다. 출수일수, 간장, 수장, 종실경도와 종실무게를 기반으로 주성분분석(PCA, Principal Component Analysis)은 stats 패키지의 prcomp () 함수로 수행하였고, 최적 군집수(K=3)는 factoextra 패키지의 fviz_nbclust () 함수로 결정하였다. 군집분석은 kmeans () 함수로 수행하였으며, 군집간 차이는 agricolae 패키지의 Tukey HSD 검정으로 평가하였다. 주요 유전자를 선별하기 위해 randomForest 패키지를 이용해 랜덤 포레스트 분석을 수행하였다. 총 500개의 트리를 구성하였고, 변수 중요도는 MDA (Mean Decrease Accuracy)와 MDG (Mean Decrease Gini)를 기준으로 평가하였다.
결과 및 고찰
국산 밀 유전자원의 농업형질 변이
국산 밀 유전자원 515점(품종 49점, 육성계통 123점, 재래종 343점)의 농업형질 및 종실특성을 2년간 전작조건에서 조사하여 분산분석을 실시한 결과, 간장, 출수일수, 수장에서 유전형(G), 환경(E), 유전형×환경 상호작용(GEI) 모두 통계적으로 높은 유의성을 나타냈다(
p<0.001). 반면, SKCS로 측정한 종실경도와 무게는 300립의 평균값을 분석에 활용한 결과, 개체간 분산이 반영되지 않아 유전형, 환경, 유전형×환경 상호작용에 대한 통계적 유의성을 검정할 수 없었다(
Table 1). 분산성분 기여율 분석에서는 모든 형질에서 유전형 효과가 가장 크게 나타났다. 출수일수(82.6%), 종실경도(85.0%), 종실무게(90.8%)에서 80% 이상의 높은 유전형 효과가 나타났고, 간장(66.5%)과 수장(59.8%)에서도 유전형 효과가 가장 높았다. 환경 효과는 출수일수(10.7%)를 제외하고는 모든 형질에서 10% 미만으로 낮게 나타났으며, 유전형×환경 상호작용은 수장(28.9%)과 간장(25.2%)에서 상대적으로 높게 나타났다.
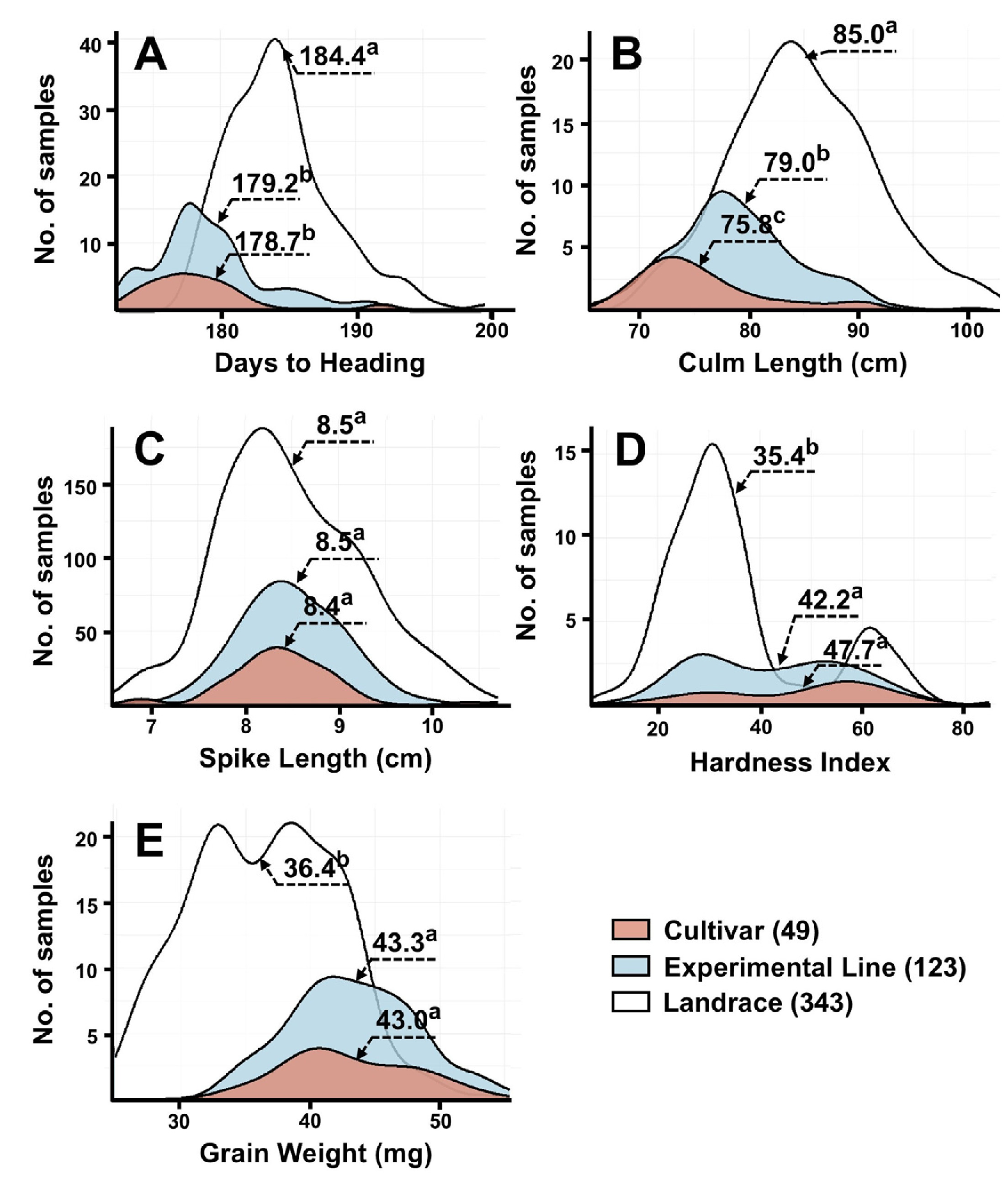
출수일수는 174-202일로 분포하였으며, 평균은 185일로 나타났다. 2022년의 평균 출수일수는 184일, 2023년은 181일이었으며, 품종 및 계통의 평균 출수일수는 179일로 재래종(185일)보다 빨랐다(
Fig. 1,
Table 2). 조품, 조한, 수원281호, 수원283호, 수원285호와 익산374호는 출수일수가 172일로 가장 짧았고, 재래종인 WIR26078과 IT166452는 201일로 가장 늦었다(Supplementary Table 2). 간장은 63.8-101.4 cm 범위였으며, 평균 81.0 cm로 나타났다. 2022년과 2023년의 평균 간장은 각각 81.0 cm와 84.4 cm였으며, 품종의 평균 간장은 75.8 cm로 가장 짧고, 재래종은 평균 85.0 cm로 가장 길었다. 조품, 조중, 탑동, 수원277호, 익산311호, 익산339호, 익산362호는 간장이 60 cm 이하로 짧았으며, 밀양25호, IT116252, IT159775, IT166442, ATRI16998, IT166431, IT166432, IT13880, IT166437는 110 cm 이상으로 길었다. 수장은 6.6-10.8 cm 범위였고, 평균은 8.6 cm였다. 2022년과 2023년의 평균은 각각 8.6 cm와 8.4 cm였으며, 품종, 계통, 재래종간 유의한 차이는 나타나지 않았다. IT140811은 5.0 cm보다 짧았고, 태중, 익산375호, IT159756, IT166420, IT166421, IT166453와 IT116254는 12.0 cm 이상으로 길었다. 종실경도와 종실무게는 각각 2.2-79.5 및 26.2-56.6 mg의 범위를 나타냈으며, 평균은 각각 33.0 및39.9 mg이었다. 2022년에는 평균 33.0 및 39.9 mg, 2023년에는 43.4 및 37.4 mg이었다. 품종과 계통은 전반적으로 재래종보다 높은 종실경도와 종실무게를 나타냈다. 수원243호, ATRI16937, HTRI16927, IT140816, IT166453, IT246976와 IT140765는 종실경도가 10 이하로 낮았고, 신미찰1호, 익산361호, 서육28호, ATRI16990, ATRI16933와 HTRI4983는 75 이상으로 매우 높았다. 종실무게는IT140797, IT140800, IT140801, IT159765, IT166448와 IT166449에서 25 mg 이하로 낮았고, 한백, 익산352호, 익산365호와 HTRI4978는 55 mg 이상으로 높았다.
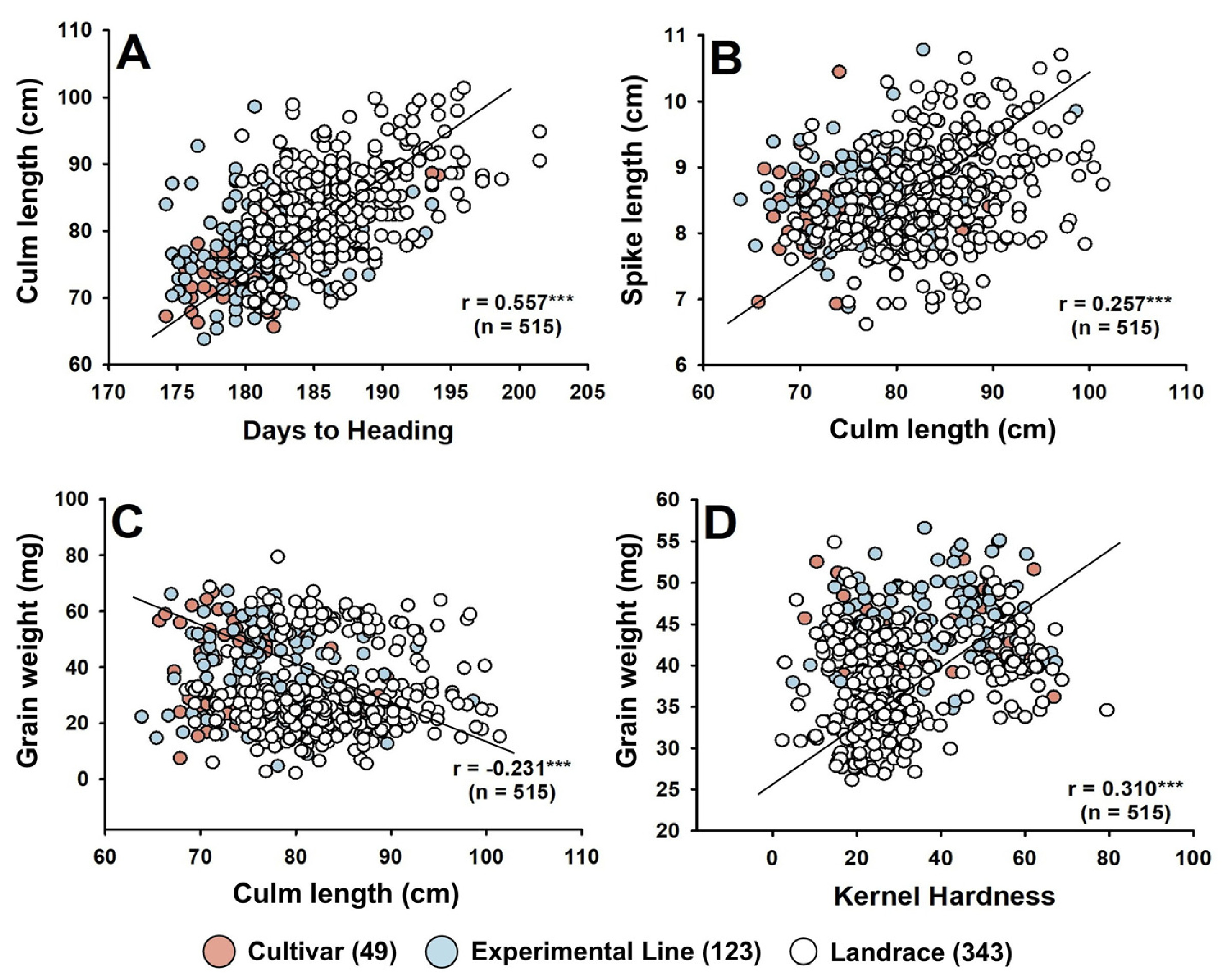
간장은 출수일수(
r=0.557,
p<0.001) 및 수장(
r=0.257,
p<0.001)와 정의 상관관계를 보였고, 종실무게(
r=-0.231,
p<0.001)는 부의 상관관계를 보였으며, 종실무게는 종실경도(
r=0.310,
p<0.001)와 정의 상관을 보였다(
Fig. 2). 국내외 품종에서 모두 간장과 수장은 양의 상관을 나타내었으며, 국내 품종에서는 간장은 천립중과 양의 상관을 보였고, 출수일수는 종실 길이와 부의 상관을 나타내었다(
Guo et al. 2018,
Shin et al. 2013,
Zhai et al. 2016).
국산 밀 유전자원의 주요 농업 형질 및 종실 특성과 관련된 유전자좌의 대립유전자 빈도를 분석한 결과,
Vrn-1,
Ppd-1,
Rht-1,
WAPO-A1,
Pin-D1,
TaCwi-A1,
TaGW2-6A,
TaSus2-2B에서 뚜렷한 유전적 변이가 관찰되었다(
Table 3,
Figs. 3,
4, Supplementary Table 3). 출수일수에 영향을 미치는 파성(vernalization) 및 광주기(photoperiod) 관련 유전자 분석 결과, 모든 자원이 겨울밀 특성을 지닌
vrn-A1 대립유전자와 광주기 둔감성(insensitive) 대립유전자인
Ppd-A1a를 지닌 것으로 나타났다(
Distelfeld et al. 2009,
Snape et al. 2001). 국산 밀 유전자원이 공통적으로
vrn-A1을 지녔지만, copy number variation (CNV) 분석 결과, 늦은 출수와 관련된 CNV≥2 유형이 전체의 59.8%로, 조숙형인 CNV=1 유형(40.2%)보다 더 높은 빈도를 보였다(
Diaz et al. 2012). CNV=1 대립유전자는 품종(71.4%)과 육성계통(65.9%)에서 높은 비율을 보였으나, 재래종에서는 CNV≥2가 73.5%로 가장 높았다.
Vrn-B1 유전자좌에서는 겨울밀형 대립유전자인
vrn-B1이 전체의 88.5%를 차지하였으며, 이는 품종, 육성계통, 재래종 간에 유사한 분포를 보였다(
Distelfeld et al. 2009). CNV=1 대립유전자를 지닌 자원의 출수일수는 180일로 CNV≥2을 지닌 자원에 비해 5일 빠른 것으로 나타났으며,
vrn-B1을 지닌 자원의 출수일수가
Vrn-B1a보다 2일 빠른 것으로 나타났다(
Table 4).
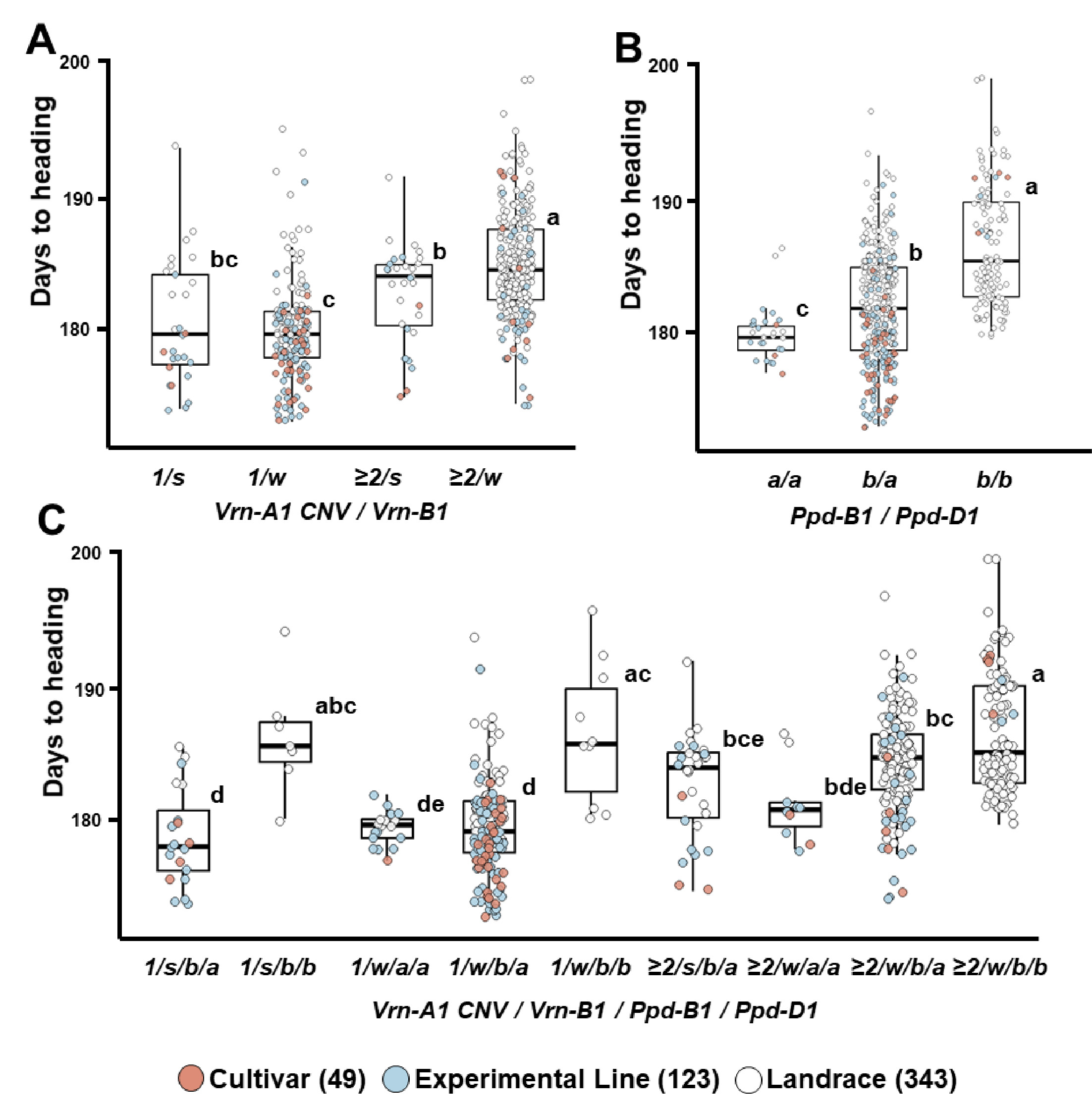
vrn-A1 대립유전자형에서 조숙형인 CNV=1을 지닌 유전자원(180일)이 CNV≥2과
vrn-B1를 지닌 유전자원(185일)에 비해 출수가 빨랐다(
Fig. 4A).
광주기 반응성에 관련된
Ppd-B1 및
Ppd-D1 대립유전자의 빈도를 보면,
Ppd-B1에서는 광주기 민감성(sensitive)인
Ppd-B1b 비율이 94.4%로 높았고,
Ppd-D1에서는 광주기 둔감성인
Ppd-D1a의 비율이 76.3%로 높았다(
Table 3). 이러한 경향은 품종과 육성계통에서
Ppd-D1a 비율이 90% 이상으로 높았으나, 재래종에서는 66.8%로 상대적으로 낮은 수준이었다. 북위 고도인 캐나다, 프랑스, 영국에서 재배되는 겨울밀은 광주기에 민감한 특성을 보이는 반면, 이탈리아 및 유고슬라비아 등 남부 지역에서는 광주기 둔감성이 뚜렷하게 나타난다. 실제로 유럽 및 북미 남부 지역에서는 광주기 둔감성 품종이 선호되는 반면, 북부 지역 품종은 민감성을 보이는 경우가 많다(
Dyck et al. 2004,
Worland et al. 1998).
Ppd-D1 유전자좌는 광주기 둔감성에 가장 큰 영향을 주며, 그 뒤를
Ppd-B1과
Ppd-A1이 따른다(
Seki et al. 2011). 또한
Ppd-D1a 대립유전자는 유럽, 캐나다, 중국의 광주기 둔감성 품종들에서 널리 발견된다(
Kamran et al. 2013,
Yang et al. 2009). 출수일수는 광주기 둔감성 대립유전자인
Ppd-B1a 및
Ppd-D1a를 지닌 자원에서 각각 3일, 4일 빨랐으며,
Ppd-B1a와
Ppd-D1a 조합을 지닌 자원은
Ppd-B1b와
Ppd-D1b 조합보다 최대 6일까지 출수일수가 단축되었다(
Fig. 4B).
Vrn-B1 유전자형에 관계없이
vrn-A1 (CNV=1)을 지닌 자원은 대부분 출수일수 179일 이하로 조숙형 특성을 나타냈다. 예외적으로, CNV=1이면서
Ppd-B1b와
Ppd-D1b를 동시에 지닌 7개 자원은 모두 재래종이었으며, 평균 출수일수가 186일로 나타나 가장 출수일수가 길었다. 이 조합에 포함된 자원 중에는 1950~1960년대에 육성된 장광, 영광, 진풍과 경광이 포함되었다. 미국 동부 연질 겨울밀과 국내 밀 품종의 출수일과
Vrn 및
Ppd 대립유전자 관계를 평가한 결과, 본 연구 결과와 마찬가지로 대부분
vrn-A1 (CNV=1)과
Ppd-D1a를 지니고 있어
vrn-A1 (CNV≥2)을 지닌 미국 밀 품종에 비해 출수일수가 매우 빨라 미국 밀 품종의 숙기 단축을 위한 자원으로 활용이 검토되고 있다(
Ma et al. 2022).
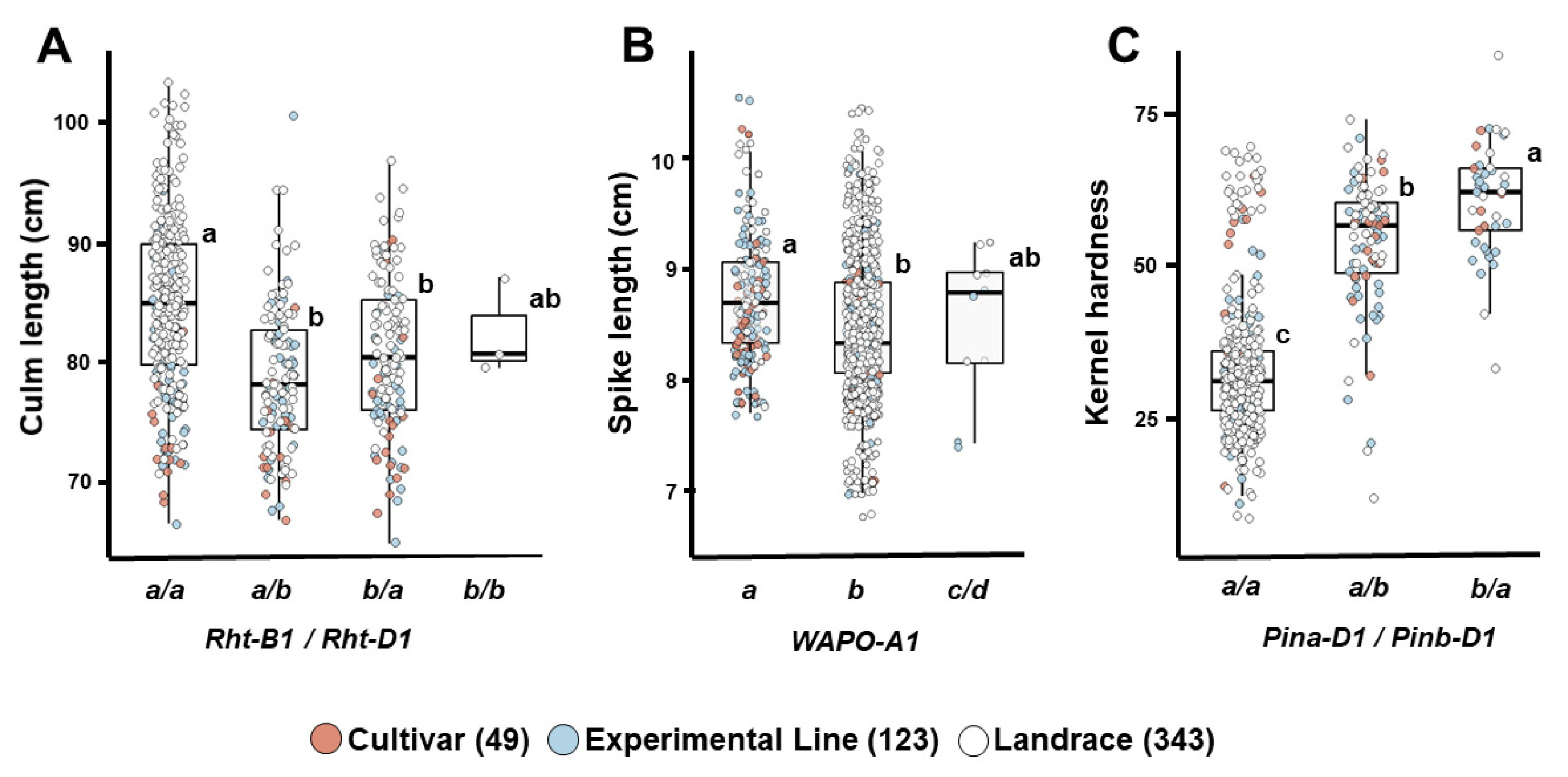
간장 감소와 관련된
Rht-B1b 및
Rht-D1b 대립유전자는 전체 국산 밀 유전자원에서 각각 24.1%와 23.3%의 빈도로 나타났다. 이들 대립유전자는 품종(36.7%, 28.6%)과 계통(27.6%, 29.3%)에서 재래종(21.0%, 20.4%)보다 높은 빈도를 보였다(
Table 3). 유전자형별 간장을 비교한 결과,
Rht-B1b와
Rht-D1b를 지닌 유전자원의 평균 간장은 각각 81.1 cm와 79.1 cm로,
Rht-B1a 및
Rht-D1a를 지닌 유전자원의 간장(83.2 cm, 83.8 cm)보다 짧았다.
Rht-B1a와
Rht-D1a 조합을 지닌 유전자원은 평균 85.0 cm로,
Rht-B1a와
Rht-D1b (79.0 cm) 또는
Rht-B1b와
Rht-D1a (81.1 cm) 조합보다 간장이 길었다. 재래종인 서육28, 서육44와 IT166460은
Rht-B1b와
Rht-D1b 조합을 지녔지만(평균 간장 82.7 cm), 이들은 다른 대립유전자 대비 간장 감소가 뚜렷하지 않았다(
Table 4,
Fig. 5A).
Rht-B1b 및
Rht-D1b 대립유전자가 간장 및 수량에 미치는 효과는 재배 환경에 따라 달라질 수 있는데, 일반적으로 간장을 약 15% 감소시켜 도복 저항성을 증가시켜서 수량을 약 24% 증가시킬 수 있다(
Flintham et al. 1997). 그러나,
Rht-D1b 대립유전자는 꽃밥 배출(anther extrusion)을 감소시켜 병원균 침입을 수월하게 해서 붉은곰팡이병(Fusarium head blight) 감수성을 증가시키고,
Rht-B1b도 붉은곰팡이병 감수성과 관련될 수 있지만 연관성은
Rht-D1b보다 낮은 것으로 알려져 있다(
Buerstmayr et al. 2020).
Rht-B1b와
Rht-D1b의 빈도는 일본 및 중국 밀에서도 유사하게 나타났으며, 중국 및 국내 재래종과 주요 품종 모두에서
Rht-B1b 빈도는 낮고
Rht-D1b는 높은 빈도를 보였다(
Cho et al. 2015b,
Yamada 1990,
Zhang et al. 2006). 중국 재래종 및 품종에서
Rht-B1b 및
Rht-D1b 빈도가 점차 증가하는 경향을 나타내며(
Zhang et al. 2006), 국산 유전자원도 2005년 이후에 육성된 국내 계통에서는
Rht-B1b 빈도는 증가하였지만,
Rht-D1b는 44.7%에서 7.1%로 급격히 감소하였다(
Cho et al. 2015a,
Cho et al. 2015b). 미국 동부 연질 겨울밀과 국내 품종의 출수일수와
Rht 대립유전자 관계 분석 결과, 미국 밀은
Rht-D1b 빈도가 56.4%로 높았으며, 국내 밀 품종은 본 연구 결과와 마찬가지로
Rht-D1a 빈도가 53.1%로 높았고,
Rht-D1b를 지닌 자원은 출수일수가 길었고,
Rht-D1a 보유 계통은 출수일수가 줄었다(
Ma et al. 2022). 본 연구에서는
Rht-B1b 및
Rht-D1b의 출수일수는 182일로
Rht-B1a 및
Rht-D1a의 183일로 차이가 없었지만,
Rht-B1b와
Rht-D1b를 지닌 자원의 출수일수는 186일로 다른 대립유전자 조성에 비해 2일 이상 길었다.
WAPO-A1 유전자는 밀의 일수립수(spikelet number per spike)를 조절하는 주요 유전자로, 이 유전자의 변이는 Promoter 영역의 115 bp 결실(P−)과 보존된 F-box 도메인 내 C47F 아미노산 치환에 따라 기능과 발현량이 달라지는 세 가지 haplotype (H1, H2, H3)으로 구분된다(
Kuzay et al. 2019). 이들 haplotype은 각각
WAPO-A1a,
WAPO-A1b,
WAPO-A1c/
d 대립유전자와 연관되며,
WAPO-A1b는 Promoter가 온전(P+)하고 C47F 변이가 존재하여 발현량이 높고 일수립수가 많아 6배체 보통밀에서 주로 발견된다. 반면, 4배체 듀럼밀에서는
WAPO-A1a가 주로 존재하고 일부
WAPO-A1c/
d도 나타난다.
WAPO-A1b는 다른 대립유전자에 비해 일수립수 및 수장 증가와 연관되며, 일부 연구에서는 수량 증가와의 관련성도 보고되었다(
Kuzay et al. 2022,
Zhang et al. 2024,
Zhou et al. 2023).
WAPO-A1은 일수립수를 조절하는 유전자이며 수장과 간접적인 연관성은 있으나, 수장의 직접적인 마커로 활용하기에는 한계가 있다. 그러나 본 연구에서는 일수립수를 조사하지 않아,
WAPO-A1을 수장 관련 대립유전자로 평가하였다. 국내 유전자원에서
WAPO-A1b의 출현 빈도는 78.6%로 높았으나, 예상과 달리 재래종에서의 비율이 92.4%로, 품종(49.0%)이나 계통(52.0%)보다 더 높게 나타났다(
Table 3,
Fig. 5B). 이러한 결과는 국내 육종 프로그램에서 일수립수 증가와 관련된
WAPO-A1 유전자에 대한 선발이 충분히 이루어졌는지에 대한 재검토가 필요함을 시사한다. 향후 국내 유전자원의 일수립수와
WAPO-A1 대립유전자 간의 관계에 대한 면밀한 분석이 요구된다. 또한, 계통 및 재래종 중 수원205호, 수원289호, 서선4호, 서선7호, HTRI4486 등 5개 자원은
WAPO-A1c/
d 대립유전자를 지닌 것으로 확인되었다. 한편,
WAPO-A1 유전자가 수장과 직접적으로 연관된 마커는 아니지만, 국내 유전자원에서는
WAPO-A1a 대립유전자를 가진 자원들이 다른 대립유전자형에 비해 상대적으로 수장이 긴 경향을 보였다.
국산 밀 유전자원의
Pin-D1 대립유전자 빈도를 보면,
Pina-D1a와
Pinb-D1a의 비율은 각각 92.0%와 82.1%로 매우 높았으며, 특히 재래종에서는 이들 대립유전자 비율이 90.1% 이상으로 압도적으로 높았다(
Table 3). 품종과 계통에서는
Pina-D1a가 82.1-85.7%,
Pinb-D1a는 60.2-68.3%로 상대적으로 낮았다.
Pina-D1a와
Pinb-D1a를 지닌 유전자원의 종실 경도는 각각 36.3과 34.9로,
Pina-D1b (60.5) 및
Pinb-D1b (53.3)에 비해 낮았으며, 이 두 대립유전자를 모두 가진 유전자원 382점의 평균 경도는 32.2로 가장 낮았다(
Table 4,
Fig. 5C).
Pina-D1b와
Pinb-D1a 유전자형을 가진 41점의 자원은 60.5로,
Pina-D1a와
Pinb-D1b (53.3)보다 경도가 높았다. 연질밀은
Pina-D1a와
Pinb-D1a를 가지며, 경질밀은 이들의 돌연변이에 의해 발생한다. 지금까지
Pina-D1에는 8종의 대립유전자(
Pina-D1b,
f,
k-n,
p,
q),
Pinb-D1에는 13종의 염기서열 돌연변이형(
Pinb-D1b-
g,
l,
q,
t,
v,
w,
aa,
ab)과 4종의 frame shift 돌연변이형(
Pinb-D1p,
r,
s,
u)이 보고되었다(
Morris & Bhave 2008). 일반적인 경질밀은
Pina-D1b 또는
Pinb-D1b를 가지며, 국내 유전자원에서는 이외의 돌연변이형은 발견되지 않았다. 같은 경질밀이라도
Pina-D1b와
Pinb-D1a 유전자형을 지닌 자원의 종실 경도가
Pina-D1a와
Pinb-D1b보다 다소 높았으며, 제분 시에는
Pina-D1b와
Pinb-D1a 유전자형이 더 많은 에너지가 요구되어 밀가루 입자가 커지고 손상 전분 함량도 증가한다(
Ma et al. 2017,
Matus-Cádiz et al. 2008,
Wen et al. 2024).
Pina-D1b와
Pinb-D1b 유전자형을 지닌 이중 돌연변이체는 단일 돌연변이체보다 종실 경도, 입자 크기 및 손상 전분 함량이 증가하였으나, 제빵 적성은 차이가 없었다. 초연질(super soft) 밀이나
puroindoline 발현이 없는 계통(
Pin null)의 개발과 관련 유전체 연구도 활발히 진행되고 있다(
Boehm et al. 2017,
Boehm et al. 2018,
Heo et al. 2013,
Martin et al. 2017).
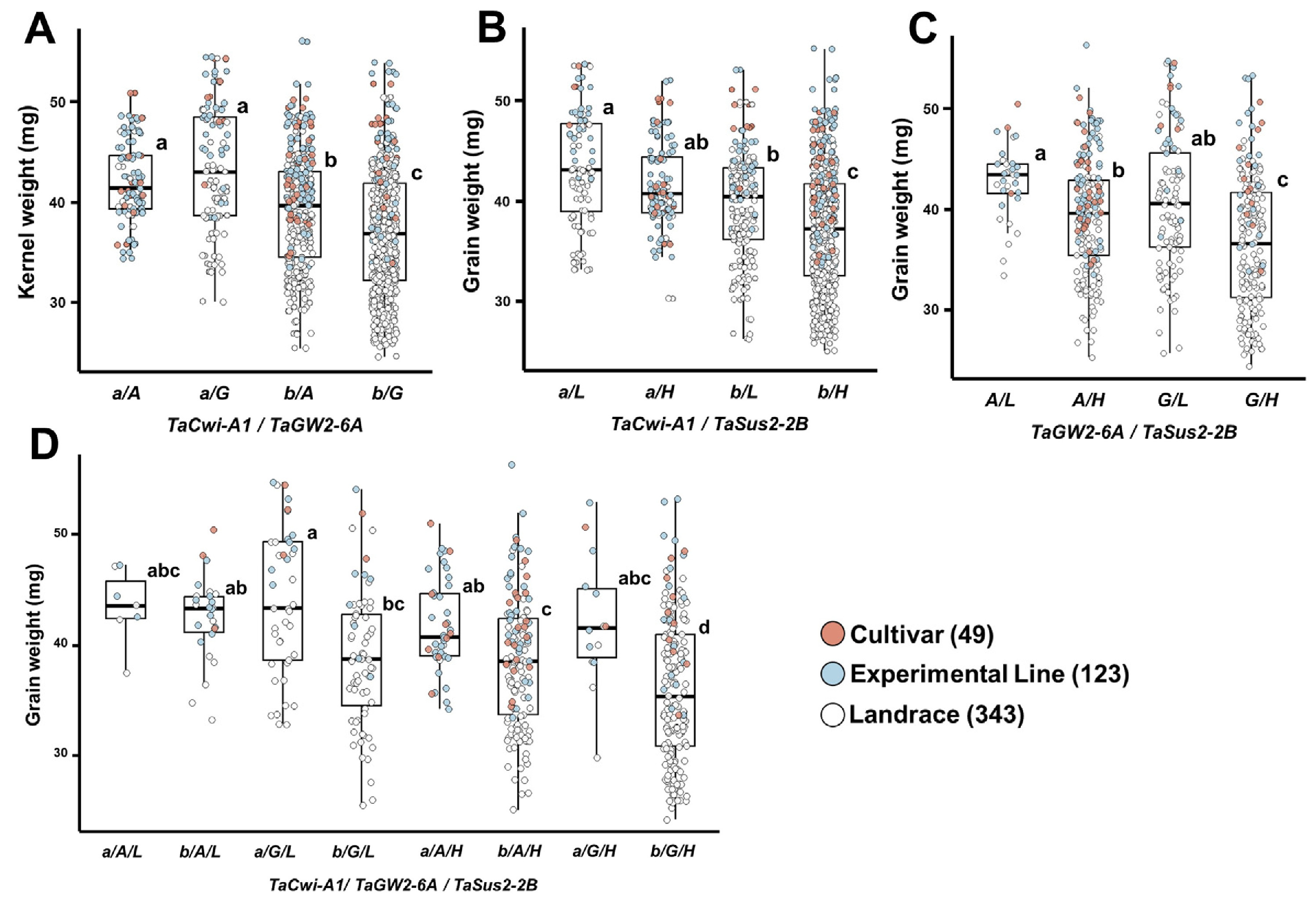
TaCwi-A1,
TaGW2-6A,
TaSus2-2B는 종실무게와 관련된 주요 유전자로, 각각 세포벽전환효소(cell wall invertase), RING-type E3 유비퀴틴 리가아제와 자당합성효소(sucrose synthase)를 암호화를 통하여 당 대사 및 종실 등숙에 관여한다(
Huo et al. 2014,
Ma et al. 2012,
Yang et al. 2012). 이들 유전자의 대립유전자 분포를 보면,
TaCwi-A1b와
TaSus2-2B-H의 빈도는 각각 79.8%와 72.6%로 높았으며, 품종, 계통, 재래종에서 유사한 경향을 나타냈다(
Table 3).
TaGW2-6A는 전체적으로
TaGW2-6A-G의 빈도(58.1%)가 높았으나, 품종 및 계통에서는
TaGW2-6A-A의 비율이 65% 이상, 재래종에서는
TaGW2-6A-G가 70.0%로 높게 나타났다. 대립유전자별 종실무게 비교에서는
TaCwi-A1b (37.7 mg),
TaSus2-2B-H (37.8 mg),
TaGW2-6A-A (39.7 mg)가 각각 다른 대립유전자보다 높았으며(
Table 4), 조합 분석에서도
TaCwi-A1a를 지닌 자원이 타 조합에 비해 종실 무게가 높았다(
Figs. 6A,
6B).
TaGW2-6A-A와
TaSus2-2B-L을 함께 가진 조합은 42.5 mg으로 가장 높았고, 반대로
TaCwi-A1b,
TaGW2-6A-G와
TaSus2-2B-H를 지닌 조합은 36.1 mg으로 가장 낮았다(
Figs. 6C,
6D). 국내 품종에서
TaCwi-A1은 천립중과 큰 차이를 보이지 않았으나, 백립계 품종에서는
TaCwi-A1a가 종자 길이와 두께, 천립중에서 우세하였다(
Kim et al. 2014).
TaGW2-6A-A는 중국, 호주, 러시아 품종에서 많이 분포하며, 이들 지역에서는 높은 천립중과 조숙성(출수일수 단축)과 연관되어 있다(
Ma et al. 2012,
Qin et al. 2014). 국내 유전자원에서도
TaGW2-6A-A는
TaGW2-6A-G보다 출수일수가 3일가량 짧았다(181일 vs. 184일).
TaSus2-2B-H는 중국 육종 프로그램에서 높은 천립중 자원 선발에 활용되고 있으나(
Huo et al. 2014,
Jiang et al. 2010), 국내 유전자원에서는 오히려
TaSus2-2B-L이 더 높은 종실 무게를 보여 기존 보고와 상반된 결과를 보였으며(
Kim et al. 2020), 이에 대한 추가 연구가 필요하다.
농업형질과 관련 대립유전자형에 따른 국내 유전자원 특성
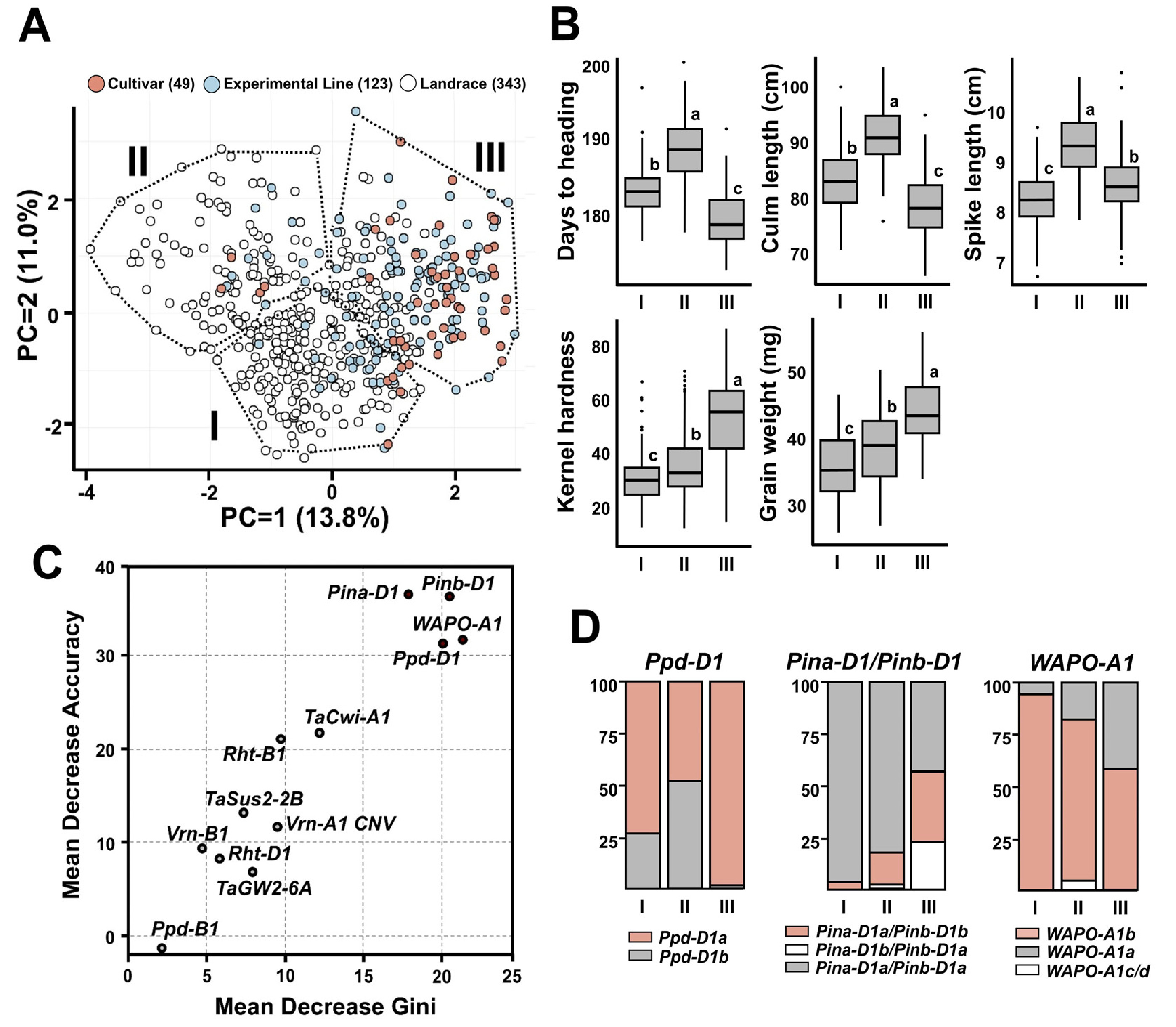
출수일수, 간장, 수장, 종실경도와 종실무게에 기반한 주성분분석(PCA) 결과, 제1주성분(PC1)과 제2주성분(PC2)은 각각 13.8%와 11.0%의 변이를 설명하였으며, 전체 자원은 세 개의 군집으로 구분되었다(
Fig. 7A). 품종과 계통은 주로 군집 III에 분포하였으며, 각각 81.6%, 68.3%의 비율을 차지하였다. 군집 III은 조숙(179.0일), 단간(78.0 cm), 높은 종실경도(50.9) 및 종실무게(43.5 mg)의 형질을 보여 국내 환경에 적합한 우수한 농업형질을 나타냈다. 재래종은 군집 I (55.1%)과 II (29.2%)에 주로 분포하였으며, 군집 I은 낮은 종실경도(29.3)와 종실무게(35.1 mg), 군집 II는 늦은 출수(188.1일)와 긴 간장(90.8 cm)으로 국내 재배에 불리한 형질을 보였다(
Fig. 7B).
군집 분류에 기여한 핵심 유전자는
Ppd-D1,
Pin-D1과
WAPO-A1로, 랜덤 포레스트 분석을 통해 도출되었다(
Fig. 7C). 이들 유전자는 평균정확도 감소값(mean decrease accuracy, MDA)와 평균 지니 감소값(mean decrease gini, MDG) 지표 모두에서 높은 중요도를 보였다. MDA와 MDG변수 중요도 평가를 위한 신뢰도 높은 지표로, 최근 밀 유전자형 분류 연구에서 널리 활용되고 있다(
Hendriks et al. 2022,
Pang et al. 2022). 이들 유전자의 대립유전자 분포는 국내 밀 육종에서 작용한 선발 압력이 뚜렷하게 나타났다(
Fig. 7D). 광주기 둔감성과 관련있으면서 조숙 품종에서 주로 나타나는 유전자인
Ppd-D1a는 품종⋅계통 중심의 군집 III에서 98.3%로 거의 고정되었지만, 재래종 중심의 군집 I (25.7%)과 II (47.8%)에서는 현저히 낮은 빈도를 나타냈다. 경질밀 조합에서 나타나는
Pina-D1a/
Pinb-D1b 또는
Pina-D1b/
Pinb-D1a)의 빈도 또한 군집 III에서 57.9%로 가장 높았고, 군집 I (3.2%) 및 II (20.0%)에서는 낮게 나타났다. 이러한 결과는 국내 육종프로그램에서 조숙과 경질 특성으로 선발이 이루어졌다고 볼 수 있다.
WAPO-A1 유전자의 대립유전자 분석 결과, 일수립수 증가와 관련된
WAPO-A1b는 재래종이 다수 포함된 군집 I (94.6%)과 II (79.1%)에서 높은 빈도로 나타났지만, 군집 III에서는 58.4%로 상대적으로 낮은 빈도를 보였다.
WAPO-A1b는 수장 증가와 관련이 있으며, 수장 증가는 일수립수 증가와 밀접한 상관을 보이지만, 종실 무게 감소가 나타나는 수량 구성 요소 간의 상쇄 관계(trade-off)가 나타날 수 있다(
Kuzay et al. 2022,
Quintero et al. 2018,
Slafer et al. 2014,
Vicentin et al. 2024). 국내 주요 밀 재배 지역은 등숙기 고온에 따른 등숙 기간 단축 현상이 뚜렷하여, 천립중이 상대적으로 높아지기 쉬운 환경이다. 국내 육종 프로그램에서는 수량 증대를 위한 주요 선발 지표로 높은 천립중이 우선적으로 고려되어 왔으며, 그 결과
WAPO-A1b의 빈도가 감소한 것으로 보인다.
천립중의 증가는 일반적으로 수량 증가와 밀접한 관련이 있기때문에, 국내 육종에서는 일수립수보다는 천립중이 높은 계통이 선발되었고, 천립중이 높아질수록 수발아율도 함께 증가하였기 때문에 생산 안정성 측면에서 주의가 필요하다(
Shin et al. 2013). 제분율 향상 및 제빵 적성 개선이라는 품질 육종 목표를 달성하기 위해, 제분율이 높고 가공 적성이 우수한 품종인 ‘금강’이 주요 교배모본으로 활용되어 왔으며, 그 결과 2000년 이후에는 천립중이 45 g 이상인 고중량 품종이 다수 육성되었다(
Choi et al. 2022). 전분이 충분히 축적된 상태에서 천립중이 증가하면 제분율과 제빵 적성이 모두 양호하게 나타나지만, 전분 축적이 부족하고 단백질 함량이 과도하게 높아질 경우 제분율은 유지되더라도 제빵 적성은 저하될 수 있다. 특히, 등숙기 고온은 글루텐 구조 형성을 방해하여 기공 형성에 부정적 영향을 줄 수 있다(
Bai et al. 2025,
Djanaguiraman et al. 2020,
Wang et al. 2018). 국내 밀 육종 프로그램에서는 수량 증대와 제빵 적성 향상을 동시에 달성하기 위한 적정 천립중에 대한 정밀한 육종 전략 수립이 필요하다. 아울러,
WAPO-A1b 마커의 활용이 일수립수 증가와 천립중 감소 사이의 수량 반응에 미치는 영향을 종합적으로 검토할 필요가 있다.
적요
본 연구는 국산 밀 유전자원(품종, 계통, 재래종 총 515점)의 주요 농업형질과 종실 특성을 2년간 전작 조건에서 평가하고, 이들 형질과 관련된 유전자좌의 대립유전자 변이를 분석함으로써 국내 밀 육종 프로그램의 기초 자료를 제공하고자 하였다. 출수기 관련 유전자 분석 결과, 조숙형 대립유전자인 vrn-A1 (CNV=1)은 전체 유전자원의 40.2%에서 나타났으며, 해당 자원은 CNV≥2에 비해 평균 5일 빠른 출수를 보였다. 또한, 겨울형 대립유전자 vrn-B1는 88.5% 빈도로 나타났으며, Vrn-B1a 보유 자원보다 평균 2일 출수가 빠른 경향을 나타냈다. 광주기 둔감성과 관련된 Ppd-B1a와 Ppd-D1a는 각각 5.6%, 76.3%의 빈도로 관찰되었고, 두 유전자를 모두 가진 자원은 최대 6일 빠른 출수일수를 보였다. 간장 관련 유전자인 Rht-B1b와 Rht-D1b는 각각 24.1%, 23.3%의 빈도로 나타났으며, Rht-B1a/Rht-D1b 조합은 Rht-B1a/Rht-D1a에 비해 간장이 6 cm 짧았다. 일수립수 증가 관련 WAPO-A1b는 전체의 78.6%에서 나타났으나, 유전자형 간 유의한 수장 차이는 관찰되지 않았다. 종실경도 관련 유전자 중 Pina-D1a (92.0%)와 Pinb-D1a (82.1%)의 빈도가 높았으며, 두 유전자를 모두 지닌 연질 자원의 평균 종실경도는 32.2로 가장 낮았고, Pina-D1b/Pinb-D1a 조합을 지닌 자원은 60.5로 가장 높았다. 종실무게 관련 유전자 중 TaCwi-A1b (79.8%), TaGW2-6A-G (58.1%), TaSus2-2B-H (72.6%)의 빈도가 높았으며, 이 세 유전자형을 모두 가진 조합의 평균 종실무게는 36.1 mg으로 가장 낮았다. 반면, TaGW2-6A-A와 TaSus2-2B-L 조합을 지닌 자원의 평균 종실무게는 42.5 mg으로 가장 무거웠다. 품종⋅계통과 재래종은 표현형 기반 주성분분석에서 서로 다른 군집에 분포하였으며, 품종⋅계통은 주로 군집 III (81.6%, 68.3%)에, 재래종은 군집 I (55.1%)과 II (29.2%)에 분포하였다. 군집 III은 조숙(179.0일), 단간(78.0 cm), 높은 종실경도(50.9) 및 종실무게(43.5 mg)를 나타낸 반면, 군집 I은 낮은 종실경도(29.3)와 종실무게(35.1 mg)를, 군집 II는 늦은 출수(188.1일)와 긴 간장(90.8 cm)을 보였다. Ppd-D1, Pina-D1, Pinb-D1, WAPO-A1이 랜덤 포레스트 분석을 통해 군집 분류의 핵심 유전자로 식별되었고, Ppd-D1a (98.3%)와 Pin-D1b (57.9%)은 군집 III에서 높은 빈도로 분포하였으며, WAPO-A1b은 군집 I (94.6%)과 II (79.1%)에서 빈도가 높았다.
사사
본 연구는 농촌진흥청 국립식량과학원의 공동연구사업(RS-2025-02214999)의 지원으로 수행되었습니다.
Fig. 1Distribution of five agronomic traits among 515 Korean wheat germplasms. (A) Days to heading, (B) Culm length, (C) Spike length, (D) Kernel hardness, and (E) Grain weight. Distributions are shown separately for cultivars (black), experimental lines (gray), and landraces (white). Superscript letters indicate significant differences among groups (p<0.05).
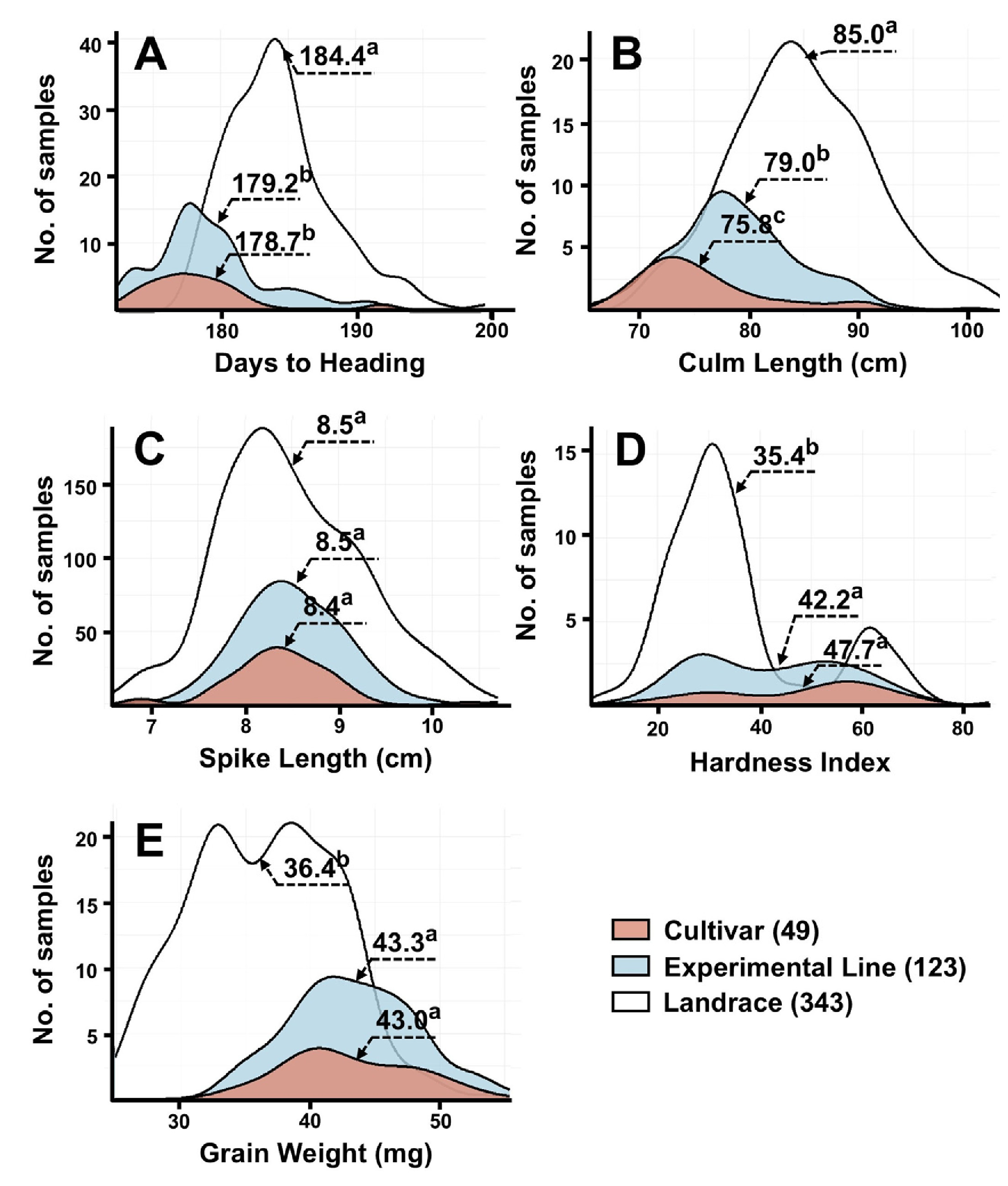
Fig. 2Relationships among key agronomic traits in 515 Korean wheat germplasms.(A) Days to heading vs. culm length, (B) Culm length vs. spike length, (C) Culm length vs. grain weight, (D) Kernel hardness vs. grain weight. Pearson’s correlation coefficients (r) and sample sizes (n) are shown in each panel.
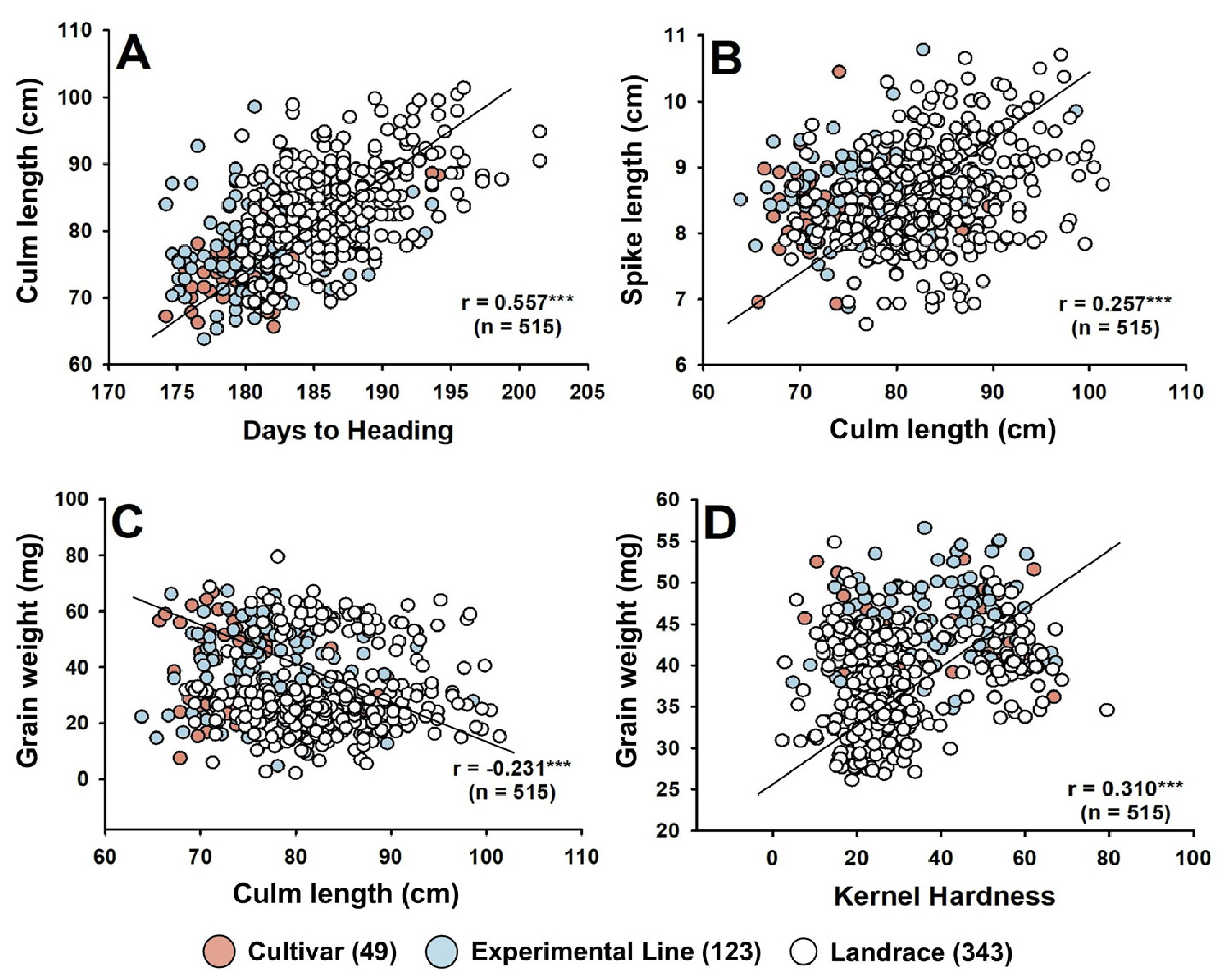
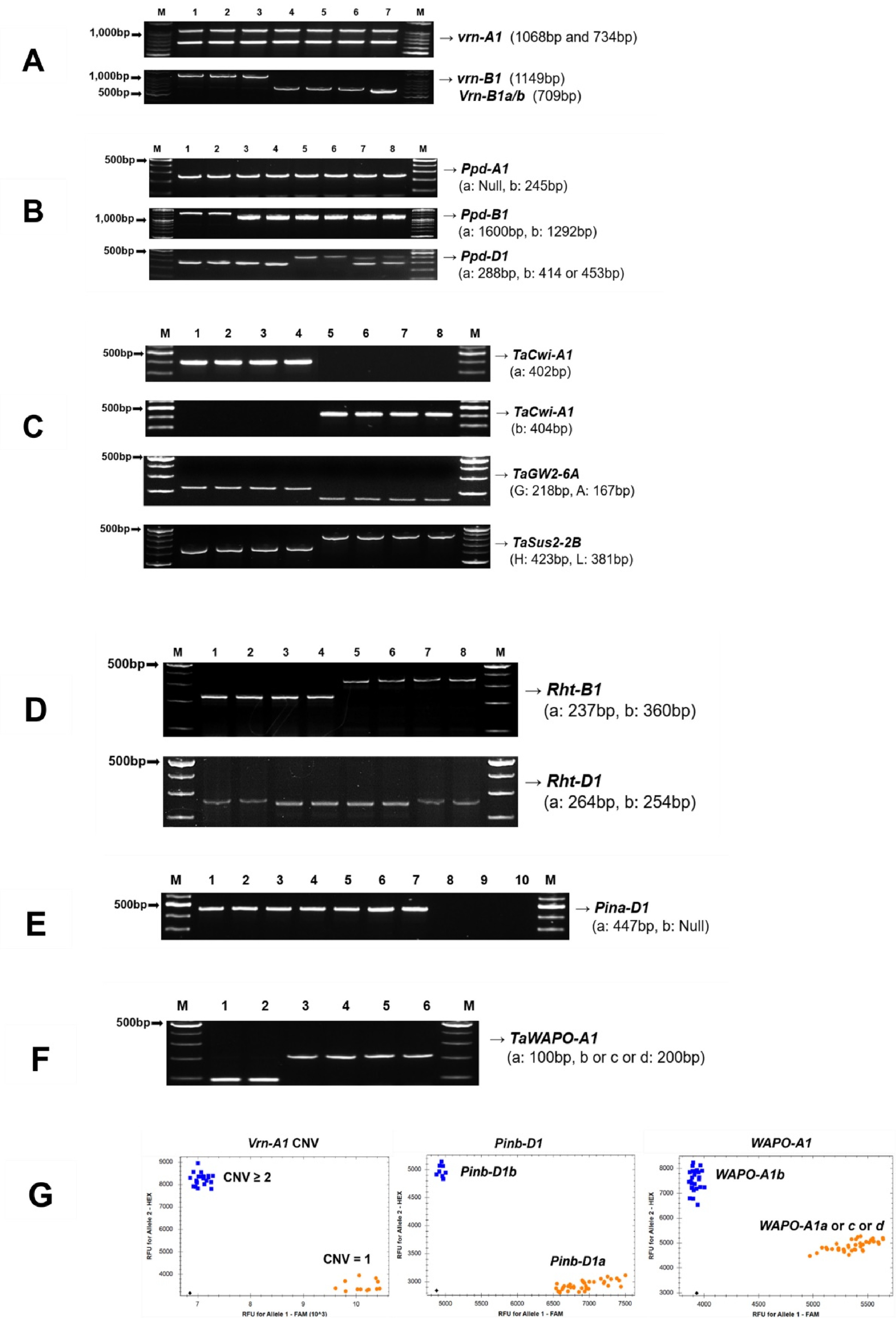
Fig. 3Amplification profiles of alleles of major genes and genotyping results in Korean wheat germplasms. (A) Vrn-A1 and Vrn-B1 (Ol, Uri, Jokyung, Joa, Iksan340, IT159748, HTRI14977); (B) Ppd-A1, Ppd-B1, and Ppd-D1 (Namhae, Alchan, Iksan313, IT151052, Chosun57, IT140777, IT140821); (C) TaCwi-A1, TaGW2-6A, and TaSus2-2B (Baekkang, Jokyung, Iksan365, IT159787, Alchan, Olgeuru, Uri, Suwon239); (D) Rht-B1 and Rht-D1 (Suan, Suwon205, Iksan311, Milyang8, Seuyuk44, IT166460, Keumkang, Suwon2599); (E) Pina-D1 (Chunggye, Suge185, Chosun30, Eunpa, Suwon92-1, IT159775, IT161214, Sukang, Suwon213, IT166464); (F) TaWAPO-A1 (Dajoong, Suge86, Suwon249, Suwon230, Seuseon7, Suwon205); (G) Genotyping clustering patterns of functional KASP markers for Vrn-A1 copy number variation (CNV), Pinb-D1, and TaWAPO-A1. M, molecular size marker. Lane numbers and corresponding germplasm names are indicated in each panel.
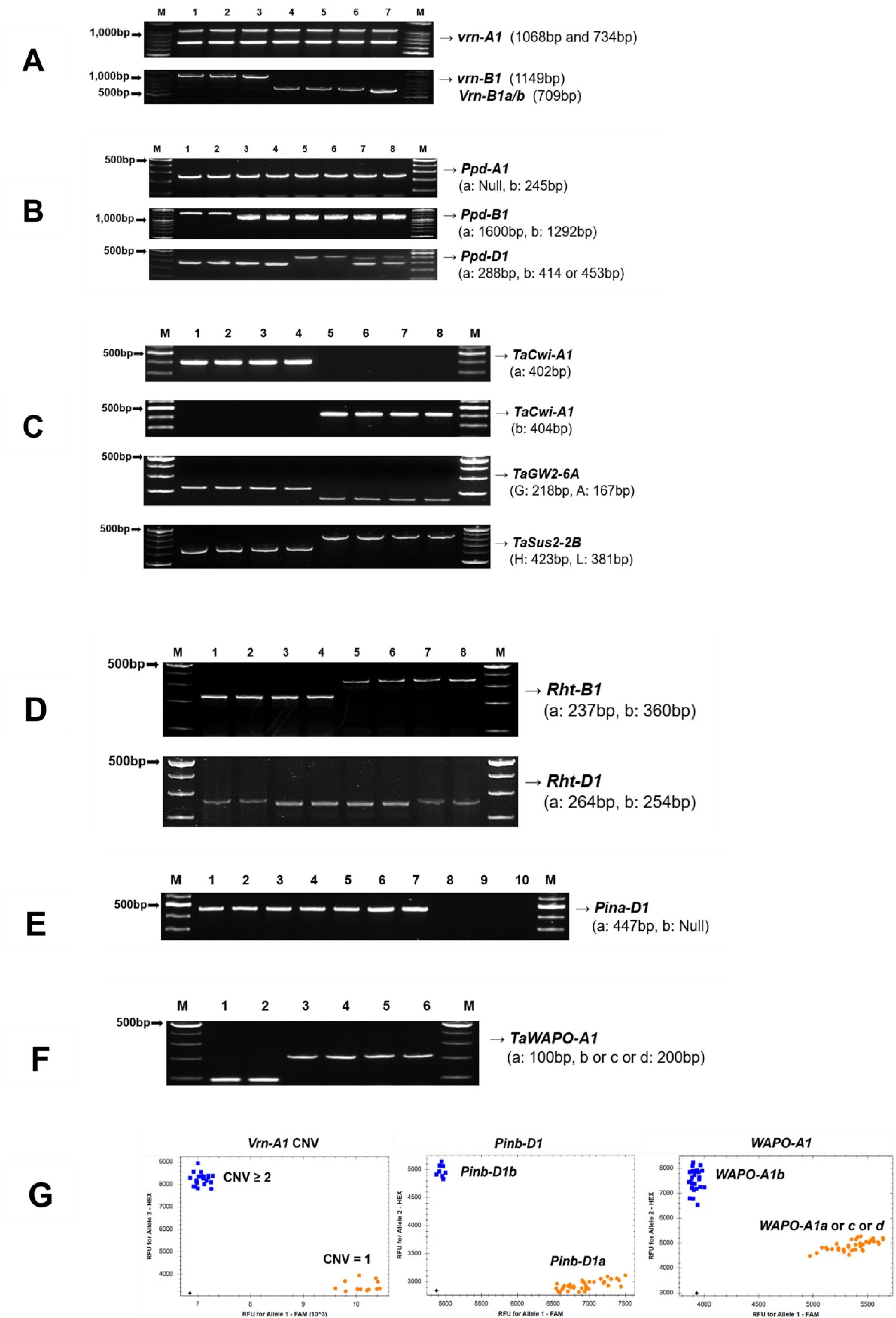
Fig. 4Differences in heading date associated with vernalization and photoperiod-related loci in 515 Korean wheat germplasms.(A) Vrn-A1 copy number variation and Vrn-B1 genotype combinations, (B) Ppd-B1 and Ppd-D1 genotype combinations, (C) Combined effects of Vrn-A1, Vrn-B1, Ppd-B1, and Ppd-D1 genotypes. At the Vrn-A1 locus, 1 and 2 indicate copy number variations; at the Vrn-B1 locus, s and w indicate spring (Vrn-B1a) and winter (vrn-B1) alleles, respectively. Different letters indicate statistically significant differences (p<0.05).
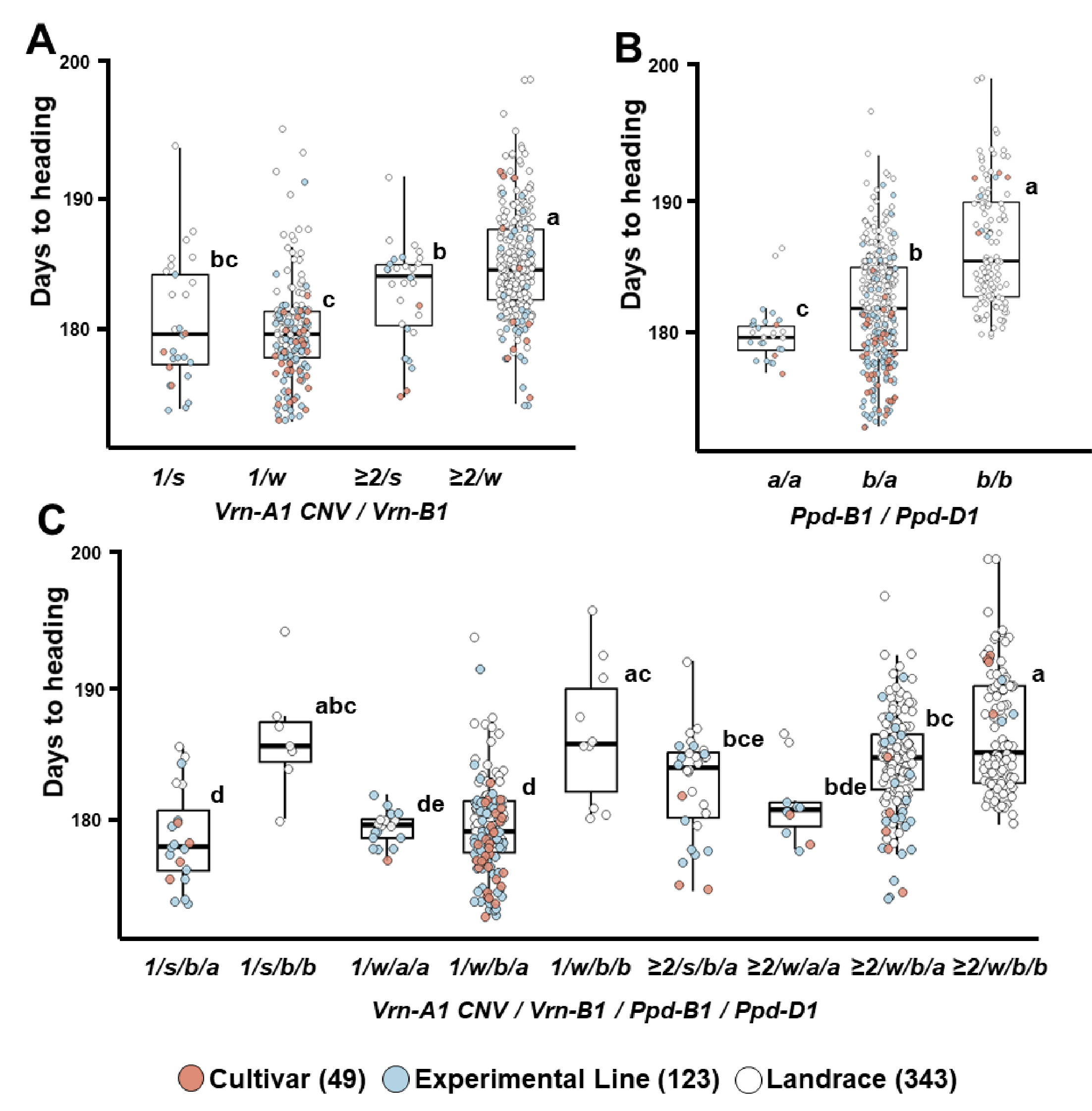
Fig. 5Differences in culm length based on combinations of Rht-B1 and Rht-D1 (A), spike length according to WAPO-A1 (B), and kernel hardness based on Pin-D1 combinations (C) in 515 Korean wheat germplasms. Different letters indicate significant differences at p<0.05.
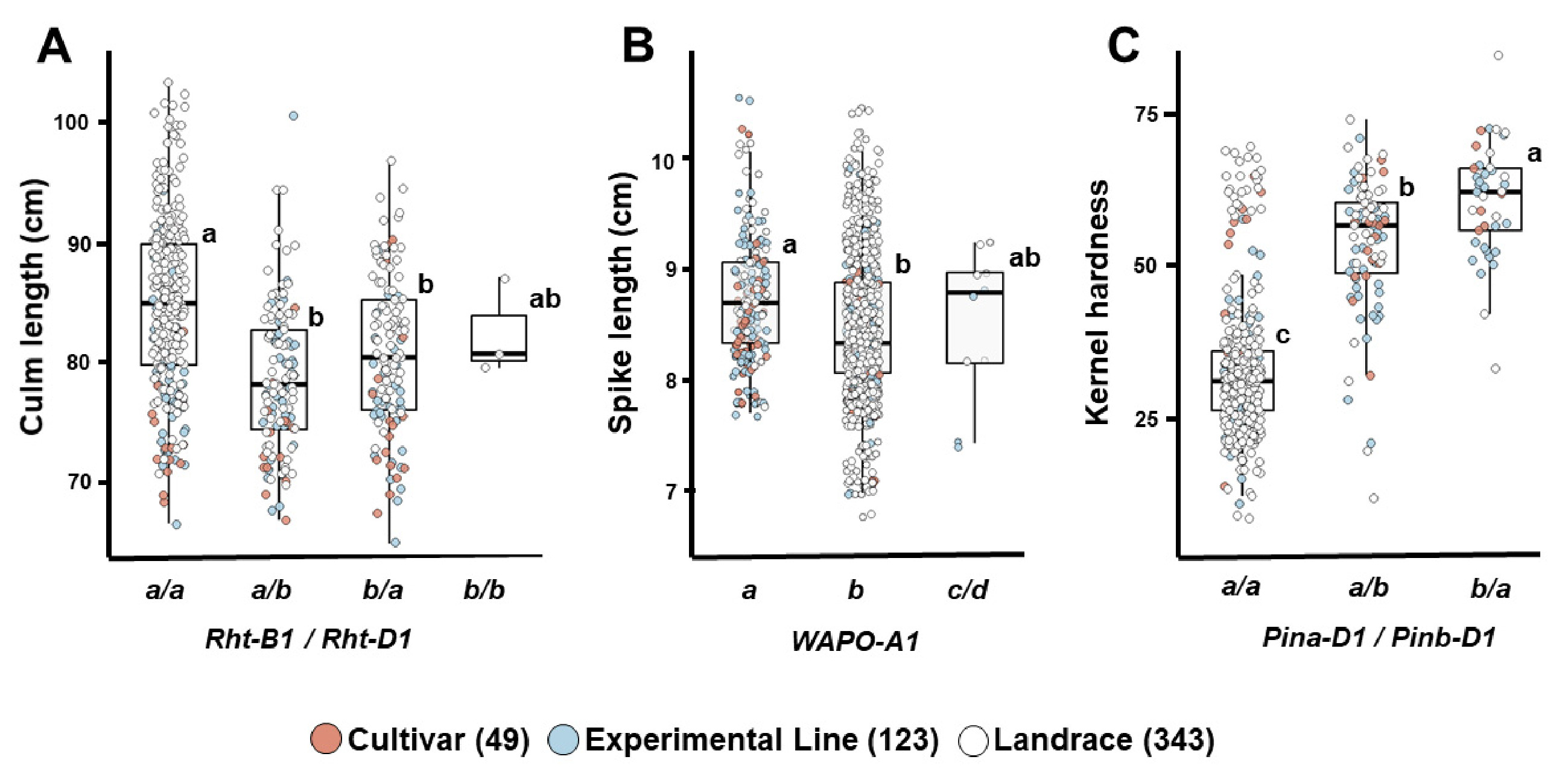
Fig. 6Differences in grain weight according to allele combinations of grain weight-related genes in 515 Korean wheat germplasms.(A) TaCwi-A1 and TaGW2-6A, (B) TaCwi-A1 and TaSus2-2B, (C) TaGW2-6A and TaSus2-2B, (D) TaCwi-A1, TaGW2-6A, and TaSus2-2B. Different letters indicate statistically significant differences (p<0.05).
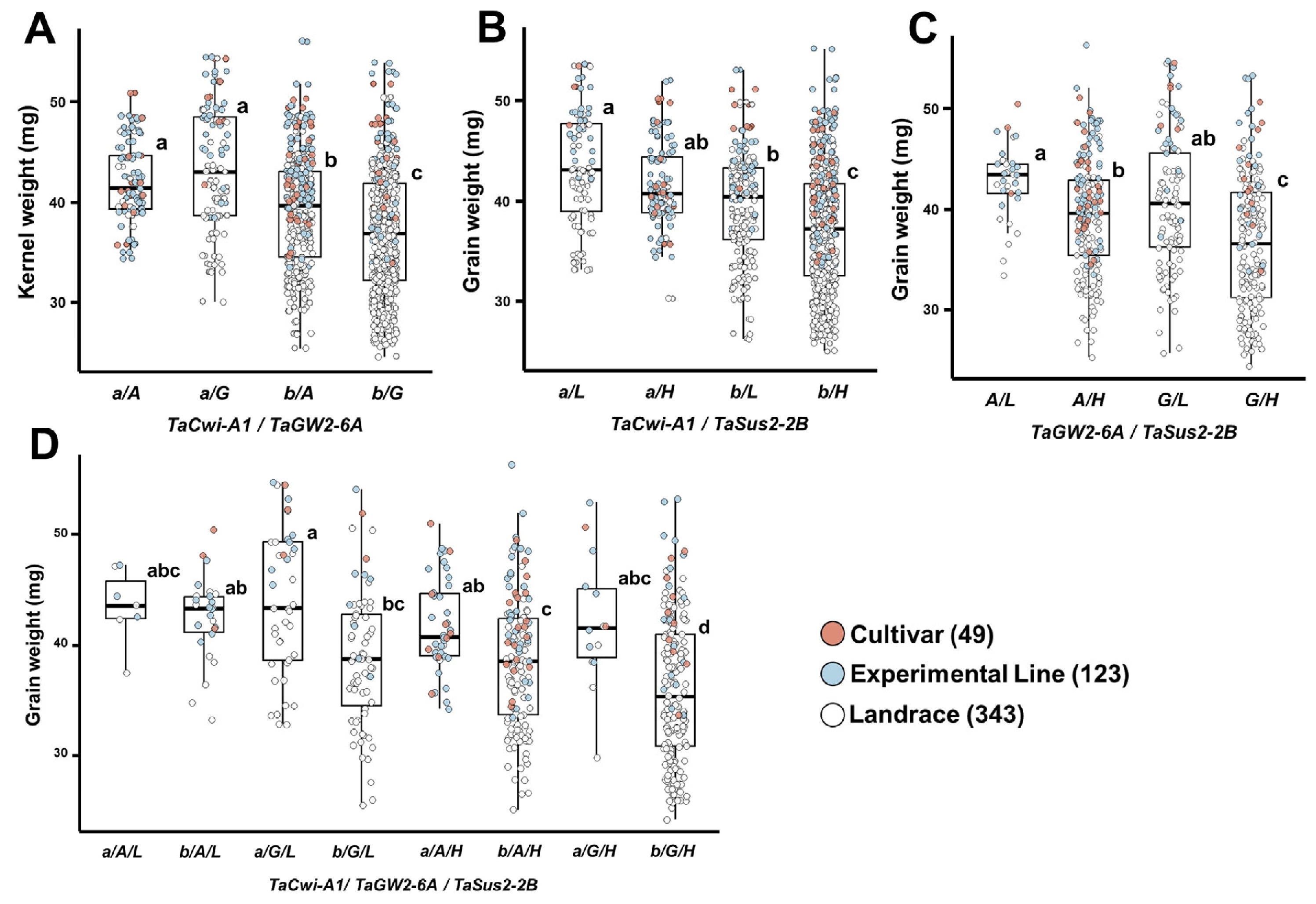
Fig. 7Phenotypic clustering and key genetic determinants in Korean wheat germplasm. (A) Principal component analysis (PCA) based on five agronomic traits. Dotted lines delineate three clusters (I-III) defined by k-means clustering. (B) Boxplots showing variation in trait values among clusters. Different letters indicate statistically significant differences (p<0.05). (C) Importance scores of genetic markers for cluster classification using the Random Forest algorithm, measured by mean decrease in accuracy (MDA) and mean decrease in Gini index (MDG). (D) Allele frequencies of the top-ranked markers from (C) across the three clusters.
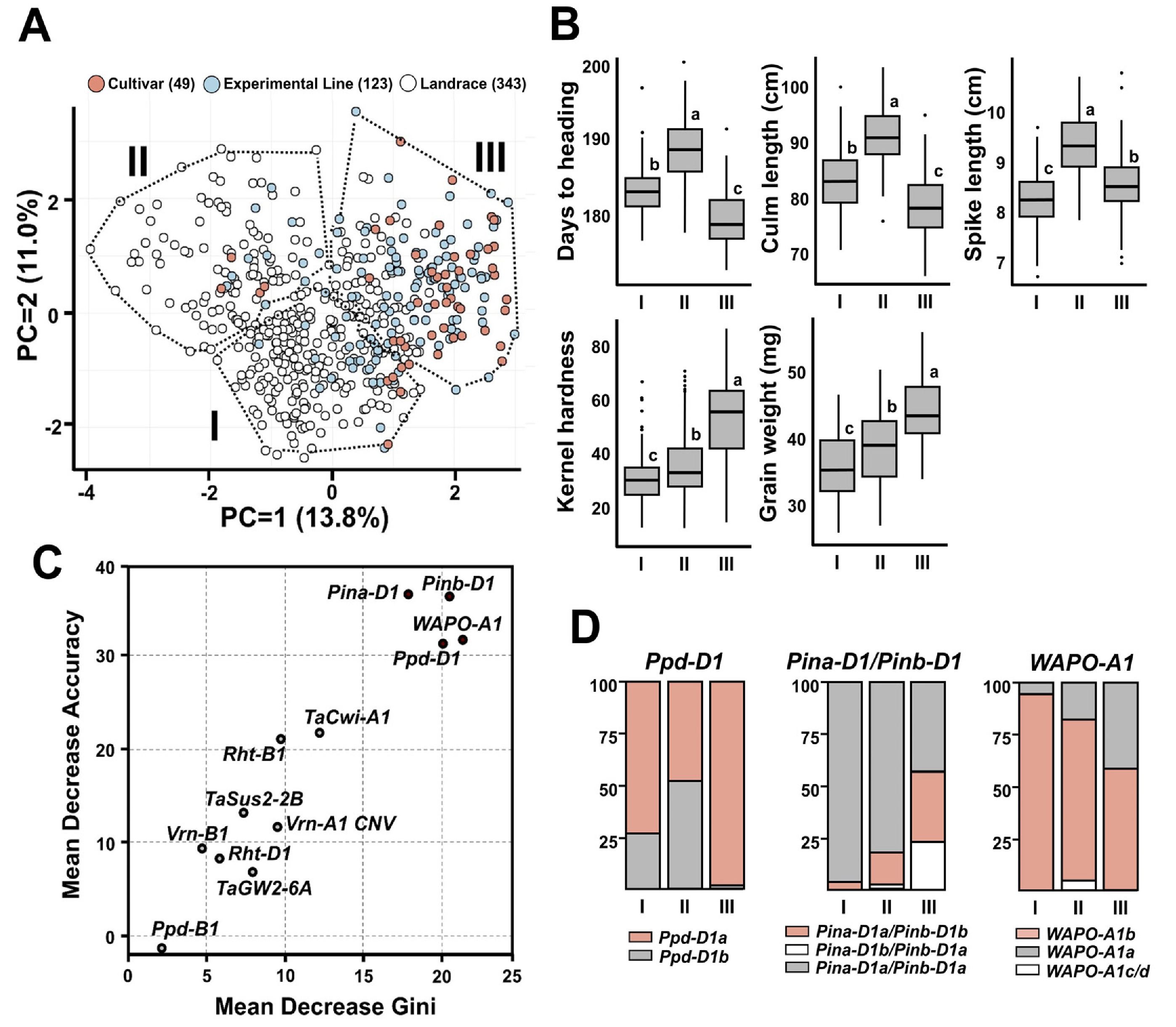
Table 1Mean squares for days to heading, culm length, spike length, kernel hardness, and grain weight in 515 Korean wheat germplasms evaluated over two years.
Table 1
|
Traits |
Mean squaresz
|
|
Variation (%)y
|
|
|
|
Genotype (G) |
Year (Y) |
G×Y |
G |
Y |
G×Y |
|
Days to heading |
159.8***x
|
10644.1***
|
12.2***
|
|
82.6 |
10.7 |
6.3 |
|
Culm length |
2647.2***
|
29135.6***
|
1011.1***
|
|
66.5 |
1.4 |
25.2 |
|
Spike length |
37.7***
|
50.8***
|
18.2***
|
|
59.8 |
0.2 |
28.9 |
|
Kernel hardness |
546.4 |
28011.5 |
42.8 |
|
85.0 |
8.5 |
6.5 |
|
Grain weight |
87.2 |
1604.3 |
5.7 |
|
90.8 |
3.3 |
5.9 |
Table 2Mean values for days to heading, culm length, spike length, kernel hardness, and grain weight in 515 Korean wheat germplasms evaluated over two years.
Table 2
|
Trait |
Germplasm |
Observation |
Year |
BLUP |
Range |
|
|
2022 |
2023 |
Days to heading
(days) |
Total |
515 |
184az
|
181a |
185±5 |
174-202 |
|
Cultivar |
49 |
180a |
176a |
181±5 |
174-194 |
|
Line |
123 |
184a |
181a |
181±4 |
174-194 |
|
Landrace |
343 |
186a |
183a |
186±4 |
180-202 |
|
Culm length
(cm) |
Total |
515 |
81.0a |
84.4a |
81.0±7.2 |
63.8-101.4 |
|
Cultivar |
49 |
70.7a |
72.5a |
74.1±5.9 |
65.7-89.6 |
|
Line |
123 |
74.8a |
78.8a |
77.3±5.9 |
63.8-98.6 |
|
Landrace |
343 |
84.7a |
88.1a |
83.3±6.6 |
68.5-101.4 |
|
Spike length
(cm) |
Total |
515 |
8.6a |
8.4a |
8.6±0.7 |
6.6-10.8 |
|
Cultivar |
49 |
8.6a |
7.9a |
8.4±0.6 |
6.9-10.5 |
|
Line |
123 |
8.6a |
8.5a |
8.6±0.6 |
6.9-10.8 |
|
Landrace |
343 |
8.5a |
8.5a |
8.6±0.8 |
6.6-10.7 |
|
Kernel
hardness |
Total |
515 |
33.0a |
43.4a |
33.0±15.3 |
2.2-79.5 |
|
Cultivar |
49 |
41.9a |
55.1a |
42.5±15.3 |
7.6-66.9 |
|
Line |
123 |
34.7a |
50.3a |
37.0±14.7 |
4.8-67.3 |
|
Landrace |
343 |
31.1a |
39.3a |
30.2±14.6 |
2.2-79.5 |
|
Grain weight
(mg) |
Total |
515 |
39.9a |
37.4a |
39.9±6.2 |
26.2-56.6 |
|
Cultivar |
49 |
45.0a |
41.6a |
44.3±4.8 |
35.2-55.0 |
|
Line |
123 |
45.7a |
41.5a |
44.5±4.8 |
34.8-56.6 |
|
Landrace |
343 |
37.1a |
35.4a |
37.7±5.5 |
26.2-54.9 |
Table 3Allele frequency at loci associated with agronomic and grain traits among 515 Korean wheat germplasm
z.
Table 3
|
Locus |
Allele |
Total (515) |
Cultivar (49) |
Experimental Line (123) |
Landrace (343) |
|
Vrn-A1
|
vrn-A1 (CNV=1)
|
207 (40.2) |
35 (71.4) |
81 (65.9) |
91 (26.5) |
|
vrn-A1 (CNV≥2) |
308 (59.8) |
14 (28.6) |
42 (34.1) |
252 (73.5) |
|
|
Vrn-B1
|
Vrn-B1a
|
59 (11.5) |
7 (14.3) |
23 (18.7) |
29 (8.5) |
|
vrn-B1
|
456 (88.5) |
42 (85.7) |
100 (81.3) |
314 (91.5) |
|
|
Ppd-A1
|
a
|
515 (100.0) |
49 (100.0) |
123 (100.0) |
343 (100.0) |
|
|
Ppd-B1
|
a
|
29 (5.6) |
3 (6.1) |
18 (14.6) |
8 (2.3) |
|
b
|
486 (94.4) |
46 (93.9) |
105 (85.4) |
335 (97.7) |
|
|
Ppd-D1
|
a
|
393 (76.3) |
45 (91.8) |
119 (96.7) |
229 (66.8) |
|
b
|
122 (23.7) |
4 (8.2) |
4 (3.3) |
114 (33.2) |
|
|
Rht-B1
|
a
|
391 (75.9) |
31 (63.3) |
89 (72.4) |
271 (79.0) |
|
b
|
124 (24.1) |
18 (36.7) |
34 (27.6) |
72 (21.0) |
|
|
Rht-D1
|
a
|
395 (76.7) |
35 (71.4) |
87 (70.7) |
273 (79.6) |
|
b
|
120 (23.3) |
14 (28.6) |
36 (29.3) |
70 (20.4) |
|
|
WAPO-A1
|
a
|
105 (20.4) |
25 (51.0) |
57 (46.3) |
23 (6.7) |
|
b
|
405 (78.6) |
24 (49.0) |
64 (52.0) |
317 (92.4) |
|
c&d
|
5 (1.0) |
0 (0.0) |
2 (1.6) |
3 (0.9) |
|
|
Pina-D1
|
a
|
474 (92.0) |
42 (85.7) |
101 (82.1) |
331 (96.5) |
|
b
|
41 (8.0) |
7 (14.3) |
22 (17.9) |
12 (3.5) |
|
|
Pinb-D1
|
a
|
423 (82.1) |
30 (61.2) |
84 (68.3) |
309 (90.1) |
|
b
|
92 (17.9) |
19 (38.8) |
39 (31.7) |
34 (9.9) |
|
|
TaCwi-A1
|
a
|
104 (20.2) |
14 (28.6) |
50 (40.7) |
40 (11.7) |
|
b
|
411 (79.8) |
35 (71.4) |
73 (59.3) |
303 (88.3) |
|
|
TaGW2-6A
|
Hap-6A-A
|
216 (41.9) |
32 (65.3) |
81 (65.9) |
103 (30.0) |
|
Hap-6A-G
|
299 (58.1) |
17 (34.7) |
42 (34.1) |
240 (70.0) |
|
|
TaSus2-2B
|
Hap-H
|
374 (72.6) |
41 (83.7) |
90 (73.2) |
243 (70.8) |
|
Hap-L
|
141 (27.4) |
8 (16.3) |
33 (26.8) |
100 (29.2) |
Table 4Differences in days to heading, culm length, spike length, kernel hardness and grain weight of 515 Korean wheat germplasmscarrying different genotypes
z.
Table 4
|
Allele |
Days to
heading |
Culm length
(cm) |
Spike length
(cm) |
Kernel
hardness |
Grain weight
(mg) |
|
Vrn-A1 (CNV=1) |
180b |
79.8b |
8.5a |
41.4a |
40.4a |
|
Vrn-A1 (CNV≥2) |
185a |
84.7a |
8.5a |
36.1b |
37.5b |
|
|
vrn-B1
|
183a |
83.0a |
8.5a |
37.9a |
38.3b |
|
Vrn-B1a
|
181b |
80.3b |
8.5a |
40.6a |
41.5a |
|
|
Ppd-B1a
|
180b |
77.2b |
8.5a |
41.7a |
39.1a |
|
Ppd-B1b
|
183a |
83.1a |
8.5a |
38.0a |
38.7a |
|
|
Ppd-D1a
|
182b |
81.2b |
8.4b |
39.8a |
40.0a |
|
Ppd-D1b
|
186a |
87.6a |
8.7a |
33.2b |
34.4b |
|
|
Rht-B1a
|
183a |
83.2a |
8.5a |
36.5b |
37.6b |
|
Rht-B1b
|
182a |
81.1b |
8.5a |
43.5a |
42.1a |
|
|
Rht-D1a
|
183a |
83.8a |
8.5a |
38.9a |
38.7a |
|
Rht-D1b
|
182b |
79.1b |
8.4a |
36.1a |
38.8a |
|
|
WAPO-A1a
|
180c |
80.7c |
8.7a |
44.1a |
44.2a |
|
WAPO-A1b
|
183b |
83.1b |
8.5b |
36.6b |
37.2b |
|
WAPO-A1c&d
|
189a |
92.3a |
8.5ab |
48.4ab |
39.5ab |
|
|
Pina-D1a
|
183a |
83.1a |
8.5a |
36.3b |
38.2b |
|
Pina-D1b
|
178b |
78.1b |
8.6a |
60.5a |
43.9a |
|
|
Pinb-D1a
|
183a |
83.1a |
8.5a |
34.9b |
37.8b |
|
Pinb-D1b
|
182a |
81.1b |
8.6a |
53.3a |
42.7a |
|
|
TaCwi-A1a
|
181b |
80.0b |
8.6a |
47.2a |
42.6a |
|
TaCwi-A1b
|
183a |
83.4a |
8.5a |
35.9b |
37.7b |
|
|
TaGW2-6A-A
|
181b |
80.2b |
8.4a |
39.2a |
39.7a |
|
TaGW2-6A-G
|
184a |
84.5a |
8.5a |
37.5a |
38.0b |
|
|
TaSus2-2B-H
|
182b |
82.3b |
8.4b |
36.9b |
37.8b |
|
TaSus2-2B-L
|
184a |
84.0a |
8.6a |
41.6a |
41.0a |
References
- 1. Bai Y, Khoddami A, Messina V, Zhang Z, Tan DKY. 2025. Response of wheat genotypes stressed by high temperature in terms of yield and protein composition across diverse environments in Australia. Agriculture 15: 514
- 2. Bentley AR, Turner AS, Gosman N, Leigh FJ, Maccaferri M, Dreisigacker S, Greenland A, Laurie DA. 2011. Frequency of photoperiod-insensitive Ppd-A1a alleles in tetraploid, hexaploid and synthetic hexaploid wheat germplasm. Plant Breed 130: 10-15.
- 3. Bettge AD, Morris CF. 2000. Relationships among grain hardness, pentosan fractions, and end-use quality of wheat. Cereal Chem 77: 241-247.
- 4. Boehm JD, Ibba MI, Kiszonas AM, See DR, Skinner DZ, Morris CF. 2017. Identification of genotyping-by-sequence tags associated with milling performance and end-use quality traits in hard red spring wheat (Triticum aestivum L.). J Cereal Sci 77: 73-83.
- 5. Boehm JD, Ibba MI, Kiszonas AM, See DR, Skinner DZ, Morris CF. 2018. Genetic analysis of kernel texture (grain hardness) in a hard red spring wheat (Triticum aestivum L.) bi-parental population. J Cereal Sci 79: 57-65.
- 6. Buerstmayr M, Steiner B, Buerstmayr H. 2020. Breeding for Fusarium head blight resistance in wheat-Progress and challenges. Plant Breed 139: 429-454.
- 7. Cho EJ, Kang CS, Jung J-U, Yoon YM, Park CS. 2015a. Allelic variation of Rht-1, Vrn-1 and Ppd-1 in Korean wheats and its effect on agronomic traits. Plant Breed Biotech 3: 129-138.
- 8. Cho EJ, Kang CS, Yoon YM, Park CS. 2015b. The effects of Rht semi-dwarfing alleles on agronomic traits in Korean wheat cultivars. Indian J Genet 76: 31-39.
- 9. Choi R, Hong S-M, Yu J-H, Lee C, Baek J, Mo Y, Park CS. 2022. Analysis of grain appearance traits and images of Korean wheat cultivars. Korean J Breed Sci 54: 158-170.
- 10. Díaz A, Zikhali M, Turner AS, Isaac P, Laurie DA. 2012. Copy number variation affecting the Photoperiod-B1 and Vernalization-A1 genes is associated with altered flowering time in wheat (Triticum aestivum). PLoS One 7: e33234.
- 11. Distelfeld A, Li C, Dubcovsky J. 2009. Regulation of flowering in temperate cereals. Curr Opin Plant Biol 12: 178-184.
- 12. Djanaguiraman M, Narayanan S, Erdayani E, Prasad PVV. 2020. Effects of high temperature stress during anthesis and grain filling periods on photosynthesis, lipids and grain yield in wheat. BMC Plant Biol 20: 268
- 13. Dyck JA, Matus-Ca'diz MA, Hucl P, Talbert L, Hunt T, Dubuc JP, Nass H, Clayton G, Dobb J, Quick J. 2004. Agronomic performance of hard red spring wheat isolines sensitive and insensitive to photoperiod. Crop Sci 44: 1976-1981.
- 14. Ellis MH, Spielmeyer W, Gale KR, Rebetzke GJ, Richards RA. 2002. "Perfect" markers for the Rht B1b and Rht-D1b dwarfing genes in wheat. Theor Appl Genet 105: 1038-1042.
- 15. Flintham E, Borner A, Worland A, Gale M. 1997. Optimizing wheat grain yield effects of Rht (gibberellin-insensitive) dwarfing genes. J Agric Sci 128: 11-25.
- 16. Fu D, Szűcs P, Yan L, Helguera M, Skinner JS, von Zitzewitz J, Hays PM, Dubcovsky J. 2005. Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol Gen Genom 273: 54-65.
- 17. Gao Y, Li Y, Xia W, Dai M, Dai Y, Wang Y, Ma H, Ma H. 2023. The regulation of grain weight in wheat. Seed Biol 2: 17
- 18. Gautier MF, Aleman ME, Guirano A, Marion D, Joudrier P. 1994. Triticum aestivum puroindolines, two basic cysteine-rich seed proteins: cDNA sequence analysis and developmental gene expression. Plant Mol Biol 25: 43-57.
- 19. Guo Z, Zhao Y, Röder M, Reif JC, Ganal MW, Chen D, Schnurbusch T. 2018. Manipulation and prediction of spike morphology traits for the improvement of grain yield in wheat. Sci Rep 8: 1-10.
- 20. Gupta PK, Langridge P, Mir RR. 2010. Marker-assisted wheat breeding: Present status and future possibilities. Mol Breed 26: 145-161.
- 21. Hendriks PW, Gurusinghe S, Ryan PR, Rebetzke GJ, Weston LA. 2022. Competitiveness of early vigour wheat (Triticum aestivum L.) genotypes is established at early growth stages. Agronomy 12: 377
- 22. Heo H, Sherman J. 2013. Identification of QTL for grain protein content and grain hardness from winter wheat for genetic improvement of spring wheat. Plant Breed Biotech 1: 347-353.
- 23. Huo J, Jiang Q, Hao C, Wang Y, Zhang H, Zhang X. 2014. Global selection on sucrose synthase haplotypes during a century of wheat breeding. Plant Physiol 164: 1918-1929.
- 24. Jiang Q, Hou J, Hao C, Wang L, Ge H, Dong Y, Zhang X. 2010. The wheat (T. aestivum) sucrose synthase 2 gene (TaSus2) active in endosperm development is associated with yield traits. Funct Integr Genomics 11: 49-61.
- 25. Jung EM, Lee DY, Kim TH, Park EJ, Song DH. 2020. Research on establishment of mid-to-long-term development plan for wheat industry in Korea. Korea Rural Economic Institute..
- 26. Kamran A, Iqbal M, Spaner D. 2014. Flowering time in wheat (Triticum aestivum L.): A key factor for global adaptability. Euphytica 197: 1-26.
- 27. Kamran A, Randhawa HS, Pozniak C, Spaner D. 2013. Phenotypic effects of the flowering gene complex in Canadian spring wheat germplasm. Crop Sci 53: 84-94.
- 28. Kim KH, Shin SH, Park S, Park JC, Kang CS, Park CS. 2014. Relationship between pre-harvest sprouting and functional markers associated with grain weight, TaSus2-2B, TaGW2-6A, and TaCwi-A1, in Korean wheat cultivars. SABRAO J Breed Genet 46: 319-328.
- 29. Kim KM, Cho SW, Choe R, Ryu NK, Hwang JY, Kim JW, Kim KH, Kang CS, Park CS. 2020. Allelic variation at loci associated with grain properties in Korean wheat cultivars. Korean J Breed Sci 52: 362-373.
- 30. Kuzay S, Lin H, Li C, Chen S, Woods DP, Zhang J, Lan T, Korff MV, Dubcovsky J. 2022. WAPO-A1 is the causal gene of the 7AL QTL for spikelet number per spike in wheat. PLoS Genet 18: e1009747.
- 31. Kuzay S, Xu Y, Zhang J, Katz A, Pearce S, Su Z, Fraser M, Anderson JA, Brown-Guedira G, DeWitt N, Peters Haugrud A, Faris JD, Akhunov E, Bai G, Dubcovsky J. 2019. Identification of a candidate gene for a QTL for spikelet number per spike on wheat chromosome arm 7AL by high-resolution genetic mapping. Theor Appl Genet 132: 2689-2705.
- 32. Lee HS, Jung J-U, Kang C-S, Heo H-Y, Park CS. 2014. Mapping of QTL for yield and its related traits in a doubled haploid population of Korean wheat. Plant Biotechnol Rep 8: 443-454.
- 33. Liu Y, He Z, Appels R, Xia X. 2012. Functional markers in wheat: Current status and future prospects. Theor Appl Genet 125: 1-10.
- 34. Ma D, Yan J, He Z, Wu L, Xia X. 2012. Characterization of a cell wall invertase gene TaCwi-A1 on common wheat chromosome 2A and development of functional markers. Mol Breed 29: 43-52.
- 35. Ma F, Brown-Guedira G, Kang M, Baik B-K. 2022. Allelic variations in phenology genes of eastern U.S. soft winter and Korean winter wheat and their associations with heading date. Plants 11: 3116
- 36. Ma X, Sajjad M, Wang J, Yang W, Sun J, Li X, Zhang A, Liu D. 2017. Diversity, distribution of Puroindoline genes and their effect on kernel hardness in a diverse panel of Chinese wheat germplasm. BMC Plant Biol 17: 158
- 37. Maccaferri M, Bruschi M, Tuberosa R. 2022. Sequence-based marker assisted selection in wheat. pp. 513-538. In: Reynolds MP, Braun HJ. (Eds) Wheat improvement. Springer, Cham, Switzerland:
- 38. Martin JM, Hogg AC, Webster RW, Giroux MJ. 2017. Creation and characterization of a double null Puroindoline genotype in spring wheat. Cereal Chem 94: 805-810.
- 39. Matus-Cádiz MA, Pozniak CJ, Hucl PJ. 2008. Puroindoline allele diversity in Canadian and northern UShard spring wheat varieties differing in kernel hardness. Can J Plant Sci 88: 807-816.
- 40. Morris CF, Bhave M. 2008. Reconciliation of D-genome puroindoline allele designations with current DNA sequence data. J Cereal Sci 48: 277-287.
- 41. Morris CF. 2002. Puroindolines: the molecular genetic basis of wheat grain hardness. Plant Mol Biol 48: 633-647.
- 42. Narasimhamoorthy B, Gill BS, Fritz AK, Nelson JC, Brown-Guedira GL. 2006. Advanced backcross QTL analysis of a hard winter wheat×synthetic wheat population. Theor Appl Genet 112: 787-796.
- 43. Pang A, Chang MWL, Chen Y. 2022. Evaluation of random forests (RF) for regional and local-scale wheat yield prediction in southeast Australia. Sensors 22: 717
- 44. Pearce S, Saville R, Vaughan SP, Chandler PM, Wilhelm EP, Sparks CA, Nadia AK, Korolev A, Boulton MI, Philips AL, Hedden P, Nicholson P, Thomas SG. 2011. Molecular characterization of Rht-1 dwarfing genes in hexaploid wheat. Plant Physiol 157: 1820-1831.
- 45. Qin L, Hao C, Hou J, Wang Y, Li T, Wang L, Ma Z, Zhang X. 2014. Homologous haplotypes, expression, genetic effects and geographic distribution of the wheat yield gene TaGW2. BMC Plant Biol 14: 107
- 46. Quintero A, Molero G, Reynolds MP, Calderini DF. 2018. Trade-off between grain weight and grain number in wheat depends on G×E interaction: A case study of an elite CIMMYT panel (CIMCOG). Eur J Agron 92: 17-29.
- 47. RDA.2012. Standard of research and analysis for agricultural technology. pp. 339-365..
- 48. Seki M, Chono M, Matsunaka H, Fujita M, Oda S, Kubo K, Kiribuchi-Otibe C, Kojima H, Nishida H, Kato K. 2011. Distribution of photoperiod-insensitive alleles Ppd-B1a and Ppd-D1a and their effect on heading time in Japan wheat cultivars. Breed Sci 61: 405-412.
- 49. Shin SH, Kim KH, Kang CS, Park JC, Hyun JN, Park CS. 2013. Effects of agronomic characteristics and grain morphology on pre-harvest sprouting in Korean wheat cultivar. Korean J Breed Sci 45: 346-357.
- 50. Shin SH, Kim KH, Son JH, Kang CS, Cheong YK, Lee CK, Park JC, Park CS. 2014. Analysis of semi-dwarf gene (Rht) construction and its relationship with agronomic characteristics, pre-harvest sprouting, and Fusarium head blight in Korean wheat cultivar. J Agric Life Sci 43: 72-79.
- 51. Slafer GA, Savin R, Sadras VO. 2014. Coarse and fine regulation of wheat yield components in response to genotype and environment. Field Crops Res 157: 71-83.
- 52. Snape JW, Butterworth K, Whitechurch E, Worland AJ. 2001. Waiting for fine times: genetics of flowering time in wheat. Euphytica 119: 185-190.
- 53. Son JH, Yang J, Kang CS, Kim KH, Kim KM, Jeong HY, Park J, Son JY, Park TI, Choi C. 2021. Annual analysis of the agronomic traits of global wheat germplasms in the Korean environment. Korean J Crop Sci 66: 120-129.
- 54. Su Z, Hao C, Wang L, Dong Y, Zhang X. 2011. Identification and development of a functional marker of TaGW2 associated with grain weight in bread wheat (Triticum aestivum L.). Theor Appl Genet 122: 211-223.
- 55. Tsilo TJ, Hareland GA, Simsek S, Chao S, Anderson JA. 2010. Genome mapping of kernel characteristics in hard red spring wheat breeding lines. Theor Appl Genet 121: 717-730.
- 56. Vicentin L, Canales J, Calderini DF. 2024. The trade-off between grain weight and grain number in wheat is explained by the overlapping of the key phases determining these major yield components. Front Plant Sci 15: 1380429.
- 57. Wang X, Hou L, Lu Y, Wu B, Gong X, Liu M, Wang J, Sun Q, Vierling E, Xu S. 2018. Metabolic adaptation of wheat grain contributes to a stable filling rate under heat stress. J Exp Bot 69: 5531-5545.
- 58. Wen Q, Lu S, Qiao P, Chen L, Hu Y. 2024. The effects of elite puroindoline gene alleles on the kernel hardness of Chinese winter wheat. Agronomy 14: 2998
- 59. Williams RM, O'Brien BL, Eagles HA, Solah VA, Jayasena V. 2008. The influences of genotype, environment, and genotype environment interaction on wheat quality. Aust J Agric Res 59: 95-111.
- 60. Worland AJ, Börner A, Korzun V, Li WM, Petrovíc S, Sayers EJ. 1998. The influence of photoperiod genes on the adaptability of European winter wheats. Euphytica 100: 385-394.
- 61. Yamada T. 1990. Classification of GA response, Rht genes and culm length in Japanese varieties and landraces of wheat. Euphytica 50: 221-239.
- 62. Yan L, Helguera M, Kato K, Fukuyama S, Sherman J, Dubcovsky J. 2004. Allelic variation at the VRN-1 promoter region in polyploidy wheat. Theor Appl Genet 109: 1677-1686.
- 63. Yang FP, Zhang XK, Xia XC, Laurie DA, Yang WX, He ZH. 2009. Distribution of photoperiod insensitive Ppd-D1a allele in Chinese wheat cultivars. Euphytica 165: 445-452.
- 64. Yang Z, Bai Z, Li X, Wang P, Wu Q, Yang L, Li L, Li X. 2012. SNP identification and allelic-specific PCR markers development for TaGW2, a gene linked to wheat kernel weight. Theor Appl Genet 125: 1057-1068.
- 65. Zhai H, Feng Z, Li J, Liu X, Xiao S, Ni Z, Sun Q. 2016. QTL analysis of spike morphological traits and plant height in winter wheat (Triticum aestivum L.) using a high-density SNP and SSR-based linkage map. Front Plant Sci 7: 1-13.
- 66. Zhang J, Burguener GF, Paraiso F, Dubcovsky J. 2024. Natural alleles of LEAFY and WAPO1 interact to regulate spikelet number per spike in wheat. Theor Appl Genet 137: 257
- 67. Zhang X, Yang S, Zhou Y, He Z, Xia X. 2006. Distribution of the Rht-B1b, Rht-D1b and Rht8 reduced height genes in autumn-sown Chinese wheats detected by molecular markers. Euphytica 152: 109-116.
- 68. Zhou J, Li W, Yang Y, Xie X, Liu J, Liu Y, Tang H, Deng M, Xu Q, Jiang Q, Chen G, Qi P, Jiang Y, Chen G, He Y, Ren Y, Tang L, Gou L, Zheng Y, Wei Y, Ma J. 2023. A promising QTL QSns. sau-MC-3D.1 likely superior to WAPO1 for the number of spikelets per spike of wheat shows no adverse effects on yield-related traits. Theor Appl Genet 136: 181