적요
Anther culture is useful and significant tool for producing haploid or doubled haploid (DH) plants in crop breeding system. Androgenesis is the way of inducing haploid and DH plants from anther (immature pollen) or microspore culture. In vitro androgenesis is efficient technique for introducing complete homozygous lines in one generation, thus less time and expense could be necessary than conventional plant breeding. In maize, anther culture is important system for shortening the breeding cycle and enhancing selection efficiency. Anther culture technique is also applicable to various researches such as molecular genetics, genetic engineering, genomics, and plant biotechnology. We review the past and present studies on anther culture and provide useful information for future researches on androgenesis in maize. The combination of androgenesis with other techniques such as molecular breeding and biotechnology is producing a variety of variety of maize species. In addition, we suggest strategy to develop androgenesis technique adapted to Korean research environment.
서 언
약배양은 화분이 들어 있는 약을 채취하여 배양하는 것으로 1964년 흰독말풀에서 어린약을 이용하여 약배양이 시작되었고, 1966년 처음으로 약배양 기술이 보고되었다(
Guha & Maheshwari 1964,
Guha & Maheshwari 1966). 한편 화분 배양은 꽃밥에 들어 있는 꽃가루를 배양하는 방법으로 약배양과 같이 반수성의 캘러스로부터 반수체 식물을 육성하여 육종에 응용되고 있는 배양 방법이다. 하지만 꽃밥에서 꽃가루를 적출하 거나 꽃가루로부터 캘러스를 유도하여 부정배를 형성시키는 것이 어려우므로 실제로는 약 전체를 배양할 수 있는 약배양을 이용하여 반수체 식물을 만든다. 이때 약(미숙화분) 및 소포자 (microspore)를 이용하여 반수체 식물을 만드는 것을 androgenesis 즉 동정생식이라고 하며, 약배양을 통해 만든 배를 androgenic embryo라고 한다. 동정생식이란 배낭 내부로 들어간 정핵이 난핵과 수정하지 않고 단독으로 분열하여 배를 형성한 것으로 약배양을 인공적인 동정생식이라고 볼 수 있다. 반면 자성생식 (gynogenesis)은 난세포에서 정상적인 수정이 없어도 세포분열 이 일어나 새로운 반수체 개체를 만드는 생식방법이다. 동정생식 과 자성생식은 작물육종가들에게 매우 흥미로운 기술이며, 공통 적으로 육종 프로그램의 시간을 절약하여 한 세대 만에 유전체 동형접합성을 달성시킬 수 있는 유용한 육종법이다(
Germanà 2011,
Portemer et al. 2015). 자성생식 연구는 1964년에 시작하 여
Tulecke (1964)에 의해 자성배우체로부터 캘러스 형성을 유도하여 처음으로 보고하였으며, 자성생식은 동정생식보다 생 식발달 및 개체 형성이 훨씬 더 느리다. 약배양 기술은 약내의 소포자들이 동정생식을 통하여 소포자 유래 배상구조 (embryo-like structure, ELS)를 유도하는 시스템이며, 배상구 조에서 바로 반수체 식물이 생성되거나 재분화가 가능한 캘러스 를 유도한 후 식물체를 육성할 수 있다. 기본적으로 약배양은 무균배양한 약으로부터 세포분열이 시작하고 캘러스(callus) 또 는 부정배가 형성하여 식물 재분화체를 얻을 수 있다. 일반적으로 기존의 계통육종법은 자가수정을 반복시킴으로써 적어도 6~7세 대가 소요되면서 유전적으로 형질이 고정된 순계인 새로운 품종 이 개발된다(
Chang & Coe 2009). 반면 약배양은 반수체 (haploid)나 반수성배(haploidy embryo)를 생산하여 육성과정 을 2~3세대 내에 완전한 순계를 개발하여 육종연한을 단축시키 고 비용도 절감할 수 있으며 유용한 열성 유전자를 가진 식물체를 선발할 수 있다(
Geiger & Gordillo 2009). 또한 반수체의 세포를 자연적 및 인위적인 염색체 배가를 통해서 동형접합성 식물체 생산이 가능하다. 특히 근친교배를 여러 번 반복하는 자화수분 작물에서 반수체의 이용은 기존의 재래방법을 대체할 수 있고 작물육종 분야에 큰 잠재력 및 발전 전망을 가지고 있다.
동정생식은 일부 작물 종에서 배양 기술 확립이 잘 구축되어 손쉽게 이용되고 있으며 약배양 및 소포자 배양을 통하여 반수성 세포에서 식물체를 유도하는 연구가 활발히 진행되고 있다(브로 콜리:
Farnham 1998, 아스파라거스:
Feng et al. 1991, 유채:
Gu et al. 2004, 배추:
Jo et al. 2012,
Seo et al. 2014 담배:
Touraev et al. 1996, 포도:
Dhekney et al. 2009, 밀:
Islam 2010a,
b). 반면 옥수수는 동정생식을 이용한 반수체 및 배가반수 체(doubled haploid, DH) 유도 및 형성이 다른 작물들에 비하여 효율성이 낮은 편이며, 약배양 및 소포자 배양은 옥수수 품종 및 유전형에 따라 영향을 많이 받기 때문에 배양 기술이 확립되어 있지 않다(
Genovesi & Collins 1982). 하지만 옥수수 육종에서 배가반수체는 육종 효율을 높이는데 중요한 배양 기술이며, 옥수 수 약배양을 이용한 반수체 배양기술은 DH 집단 생산을 위하여 효과적인 기술이다(
Mohammadi et al. 2007). 약배양은 공여식 물의 유전자형(genotype), 소포자 발달 단계, 환경요인 등에 영향을 미치며, 특히 약배양에서 약의 유도와 재분화체 생산을 위한 배지 조성 및 배양 조건, 물리적 및 화학적 조건은 매우 중요한 요인이다(
Kahrizi et al. 2000,
Spitkó et al. 2006). 따라서 약배양은 육종적 이용 가치가 높고 조직배양의 한 기술로서 다양한 품종에 적용 및 응용할 수 있으며, 돌연변이 육종과 형질 전환 기술에도 응용할 수 있는 육종기술로서 매우 유용하다 (
Karsai & Bedö 1998).
본 론
옥수수 약배양에 미치는 요인
옥수수 약배양은 중국에서
401 research group(1975)에 의해 시작하여
Kuo et al. (1978)이 다양한 옥수수 품종으로부터 약배양 효율성을 검정하여 발표하였으며, 옥수수 소포자(microspore) 배양은
Nitsch (1977)에 의해서 처음으로 보고되었다. 이후 약배 양에 적합한 옥수수 품종을 선발하고 약배양의 효율성을 높이기 위하여 다양한 연구가 시작되었다(
Dieu & Beckert 1986,
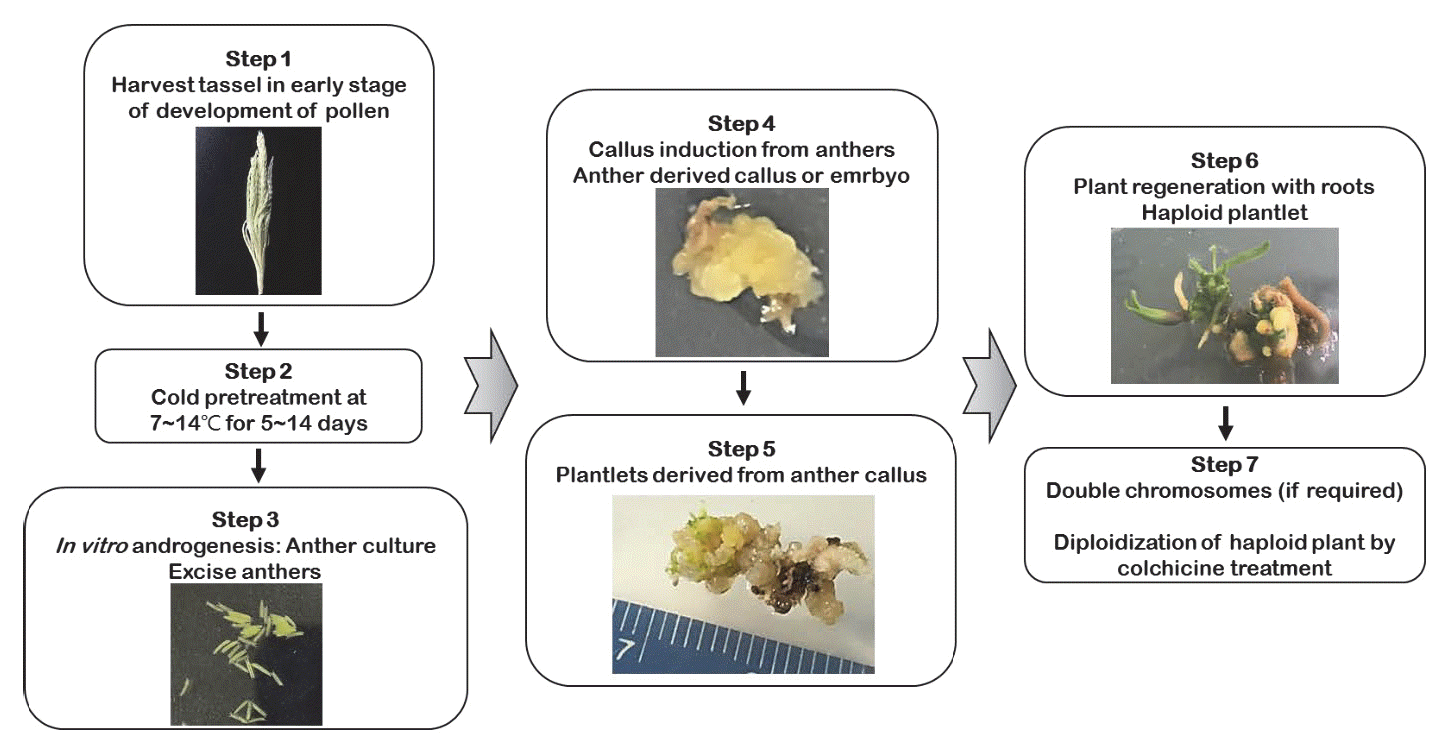
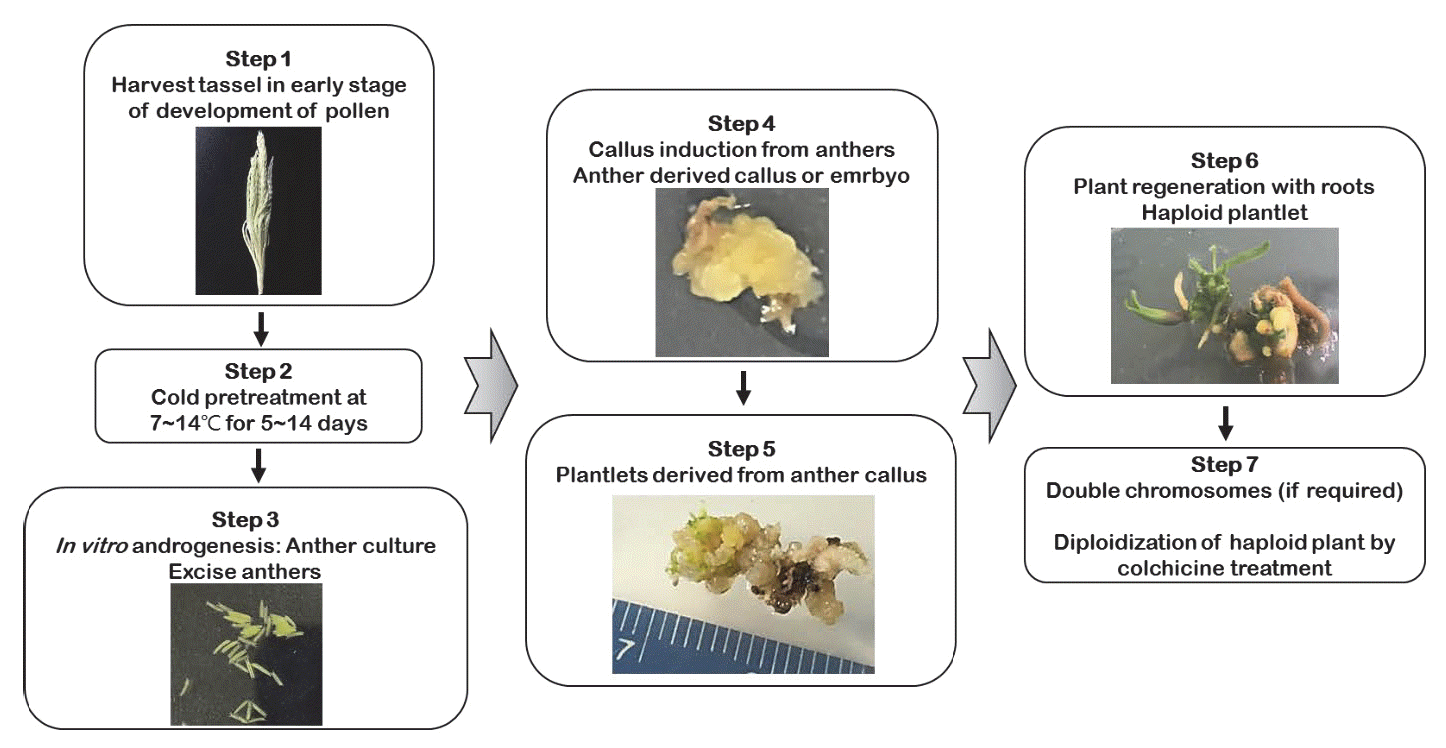
Genovesi & Collins 1982). 일반적으로 옥수수를 포함한 대부분 의 작물에서 약배양 기술 시스템은 대체로 유사한 실험과정과 방법으로 연구가 이루어지고 있으며, Fig.
1에서 약배양의 각 단계별 및 작업 과정을 확인할 수 있다. 옥수수 약배양 기술의 최대 장점은 배가반수체 식물에서 열성유전자의 동형접합성 계통 개발이 전통 육종기술보다 빠르며, 식물 육종가와 유전학자 들에게 매우 가치 있는 기술이란 점이다(
Jumpatong et al. 1996). 현재 약배양 기술은 상당히 발전하였으나, 옥수수 약배양 연구는 다른 작물에 비해 효율성이 비교적 낮기 때문에 반수체와 DH 기술을 이용하기 위하여 다양한 조건 및 요인들이 고려된다.
Fig. 1.Schematic diagram of haploid and doubled haploids selection following anther culture technique in maize.
옥수수 유전자형
염색체 배가 및 DH 생산
한편 동정생식을 통한 반수체 캘러스와 식물체는 자연적으로 염색체 복제 및 배가가 나타나는 경우는 매우 낮다(
Chase 1969). 일반적으로 약배양에서 DH 생산 및 염색체를 배가시키기 위하 여 인위적으로 colchicine(체세포분열 억제)을 주로 사용하고 있다. 하지만 colchicine은 암유발성이 매우 큰 물질이므로 취급 및 사용 후에는 폐기가 중요하다. 이러한 단점을 보완하고 colchicine을 대체하기 위하여 제초제를 사용하여 염색체 배가 연구가 진행되고 있으며,
Wan et al. (1991)은 옥수수 약배양에 서 유도된 캘러스로부터 DH 식물을 생산하기 위하여 제초제 (amiprophosmethyl/AMP, pronamide, oryzalin, trifluralin)를 반수체 캘러스에 처리하였으며, AMP와 pronamide를 처리한 실험구에서 배가배수체를 유도하는데 유용하다고 보고하였다. 실제로 제초제가 체세포분열의 효과적인 억제제로 보고되고 있으며(
Häntzschel & Weber 2010), 제초제는 colchicine에 비 하여 가격 비용이 저렴하며 독성은 거의 없고 취급이 용이하다.
약배양에서 스트레스 전처리는 약배양 과정 중 소포자 발달단 계를 촉진하는 유도인자로 매우 중요한 요인이며(
Pechan & Smykal 2001), 스트레스 전처리를 통해 평범한 꽃가루는 배 발생이 가능한 캘러스를 형성하고 이 과정은 동정생식에 있어서 중요하다(
Uváčková et al. 2012). 약배양은 웅성화 배아 (androgenic embryo)로부터 완벽한 동형접합성 반수체 및 배가 반수체 작물의 생산이 가능하다. 약배양의 효율성을 높이기 위하 여 배양 전 수꽃 및 약에 질소가스 노출, 생장호르몬 분무, 수분 부족 등의 다양한 스트레스를 가하여 약으로부터 배 발생을 유도한다. 하지만 대부분의 작물에서 약 및 소포자 배양을 위해 온도 전처리를 통하여 배양의 효율성을 증진시킨다. 배추속 식물 과 담배의 약과 소포자 배양에서 화서 및 화뢰에 32~35°C의 고온처리는 배 발생에 효과적이며(
Seo et al. 2014,
Touraev et al. 1996), 옥수수와 같은 화본과 및 단자엽 식물은 저온 처리가 더 효과적이다. 특히 옥수수는 약배양 전 수꽃에 주로 7~10°C의 저온 처리(
Genovesi & Collins 1982,
Hosseini et al. 2015,
Ismaili & Mohammadi 2016)를 통하여 약배양에 이용하고 있으며, 과거 몇몇 연구에서는 저온과 고온을 혼합 처리(
Coumans et al. 1989,
Genovesi 1990)하여 약배양의 효율 성을 높였으나 현재는 저온 처리를 주로 사용한다(Table
1). 한편 약배양 초기과정에 colchicine을 처리하여 약배양의 효율성을 높이는 것을 확인하였으며(
Barnabás et al. 1999,
Mohammadi et al. 2007,
Obert & Barnabás 2004,
Wan et al. 1989), 또 다른 연구에서는 저온 처리 및 스트레스 전처리 과정을 생략하고 다양한 배지를 조성하여 바로 약배양 과정을 거쳐 배 발생이 유도되었다 (
Barnabás et al. 1987,
Dieu & Beckert 1986,
Hassan et al. 2001,
Nitsch et al. 1982). 하지만 약배양 전에 수꽃 및 화서에 고온과 저온의 전처리 과정이 소포자의 분화 및 발생에 있어서 영향을 미치는 것에 대하여 현재까지 정확하게 밝혀진 연구가 많지 않으나, 밀 약배양 연구에서 생리학적 요인들 (약배양과 재분화 배지)과 고온 등에 의해서 알비노 빈도를 증가 시킬 수 있다고 보고되었다(
Indrianto et al. 2001,
Islam 2010a). 또한
Torp & Andersen (2009)은 알비노 식물 생산의 빈도율을 감소시키기 위해서 장기적인 저온 처리는 피하라고 제안하였으 며, 단기간(3~4일)의 저온 처리를 통하여 소포자 생존율을 증가 시켜 알비노 빈도율을 줄일 수 있었다.
Table 1.Application of pretreatment for in vitro androgenesis in maize.
옥수수 약배양 배지 조성
옥수수 약배양에서는 일반적으로 N6 (
Chu et al. 1975)와 YP (
Ku et al. 1977,
Genovesi & Collins 1982)를 기본배지로 이용하 여 식물생장조절제 조성, sucrose, 활성탄(activated charcoal), gelling agent (phyto agar, phytagel) 및 아미노산 등을 첨가하여 사용하며, 특히 sucrose는 옥수수 약배양 유도에 있어서 매우 중요하다.
Barnabás et al. (1987)은 15%의 고농도 sucrose를 첨가하여 옥수수 약배양의 효율성을 확인하였으며,
Pescitelli et al. (1990)은 다양한 sucrose의 농도(4.5~18%)를 조성하여 소포자 배양에 검증한 결과 비교적 높은 농도인 8~9.5% 사이에서 소포자 유래 배상구조(embryo-like structure, ELS)가 다른 농도 에서보다 높게 반응하였다. 대부분의 옥수수 약배양은 9%의 sucrose를 주로 사용하지만(
Büter et al. 1991,
Büter et al. 1993,
Saisingtong et al. 1996), 옥수수 유전자형에 따라 비교적 낮은 6%의 농도와 식물생장조절제의 조성으로 약배양의 배 발생 효율성이 높게 나타났다(
Hosseini et al. 2014). 최근에는 YP배 지에 6%의 sucrose 및 maltose와 0.0125 M의 polyethylene glycol (PEG)를 첨가하여 약배양의 효율성을 평가하였으며, 결과 적으로 약배양 배지에 낮은 수준의 PEG 첨가는 옥수수 약배양을 향상시킬 수 있는 요인으로 확인되었다(
Ismaili & Mohammadi 2016). 또한
Kasha et al. (1990)은 화본과 작물의 약배양 및 소포자 배양에서 maltose을 사용하면 약으로부터 배 발생 및 재분화체 유도 효율성이 높았으며, sucrose와 maltose는 품종의 유전형에 따라 배 발생을 억제하거나 촉진시키는 것으로 나타났 다. 한편 약배양 배지에 활성탄을 첨가하면 배 발생률이 증가하여 대부분의 연구자들이 약배양 및 소포자 연구에 활용하고 있다 (
Büter et al. 1993,
Büter et al. 1991,
Obert & Barnabás 2004,
Pescitelli et al. 1990,
Saisington et al. 1996,
Spitkó et al. 2006,
Wan et al. 1989). 활성탄은 배지 내에서 가스 및 고체 화합물을 흡수시키기 때문에 세포 발달에 저해될 수 있는 각종 물질들을 억제할 수 있으며, 식물생장조절제 중 auxin과 cytokinin 등이 과잉으로 작동하면 바로 흡착하여 약배양의 효율성을 높일 수 있다(
Büter et al. 1993).
분자육종은 분자생물학적 기술을 적용 및 응용 시키는 것으로 분자마커를 활용하여 유용한 작물들을 선발하는데 이용할 수 있으 며, 주로 QTL mapping, marker-assisted selection (MAS), 유전공 학 및 식물형질전환 등의 다양한 기술이 있다. 특히 약배양과 DH 기술은 옥수수 분자육종에 응용 및 활용되어 새로운 계통을 육성할 뿐만 아니라 시간과 노동 등의 절감으로 전통육종 방법보다 효과적 으로 계통 선발이 가능하다(
Prasanna et al. 2012). 또한 분자마커를 이용하여 옥수수 병저항성 및 재해내성 등 다양한 유전자들이 포함 된 형질들을 선발하여 품종 육성이 가능하며, 양친의 완벽한 동형접 합성 동형유전자형은 품종보호에 필요한 DUS (distinctness, uniformity, stability; 구별성, 균일성, 안정성) 조건에 부합하여 새로운 품종 육성 연구에 활용할 수 있다(
Geiger & Gordillo 2009,
Prasanna et al. 2012). 또한 육성된 계통들을 유지하기 위하여 투자되는 시간과 비용 및 노력 등이 경감하여 분자육종과 작물 유전학 연구를 특정한 계절에 상관없이 비교적 쉽게 수행할 수 있는 가능성이 충분하다(
Röber et al. 2005). 따라서 옥수수 육종에 있어서 DH 기술과 분자마커의 조합은 분자 육종 연구의 효율성을 증진하고 분자 유전적 결과물에 대하여 발전 가능한 전략이다 (
Prasanna et al. 2012). 또한 유전자 지도를 작성하여 표현형의 차이가 나타난 DH 집단의 양친(모계 및 부계에서 발현되는 유전자 원)을 이용할 수 있으며, 분자마커와 형질 연관 관계 분석을 통해 증명된 분자마커들을 MAS에 응용할 수 있다(
Chang & Coe 2009). 특히 유용한 유전자 및 대립형질 등을 활용하여 상대적으로 단순하 게 유전되는 형질을 갖는 DH 계통의 MAS와 seed-chipping 기술을 접목하면 직접적으로 DH 계통의 표현형을 비교 및 확인하는 것보 다 비용이 절감되고 빠르며 훨씬 효과적이다(
Prasanna et al. 2012). 최근에는 약배양으로부터 배 발생 유도에 중요한 요인인 저온 전처 리를 프로테오믹스(단백질 유전정보학) 연구에 접목해서 소포자 배발생 유도의 분자 메커니즘을 증명하여 새로운 정보를 제공하고 있다(
Uváčková et al. 2012). 이 연구에서 중요한 사실은 약배양 유도 중에 과산화 수소(hydrogen peroxide)를 분해하는 효소인 아스 코베이트 퍼록시다아제(ascorbate peroxidase)가 upregulate하였 고, 이것은 단백질 유전 정보 데이터에서 입증되어 약배양에서 동정생식이 유도되는 동안에 퍼록시다아제 활성이 증가되는 것을 확인하였다. 이러한 단백질 유전 정보 데이터는 옥수수 동정생식 유도 중에 산화성 스트레스(oxidative stress)를 조절하는 효소들에 대한 역할을 밝혀내어 새로운 정보를 제공할 수 있다.
약배양은 대량의 배(embryo)가 유도되고 동일한 유전자를 지닌 반수체 및 단세포이므로 형질전환 연구에 적용하면 유용 유전자 선발 및 새로운 품종 개발을 위하여 효율성 높은 기내 배양 기술이다. 약배양 형질전환 연구는 단 한 세대를 통하여 반수체로부터 배가된 형질전환체를 복구 및 재연이 가능하고, 이러한 형질전환체들은 이식 유전자뿐만 아니라 전체 genome 역시 모두 동형접합성이다.
Gaillard et al. (1992)은 옥수수 소포자로부터 형질전환에 적합한 소포자의 시기를 확인하기 위해 서 microinjection을 이용한 형질전환을 시도하였으며, 전제 배 발생 구조에서 초반 4~10일 정도된 소포자가 이보다 늦은 단계의 소포자보다 활력이 더 높아 형질전환에 적합하였다.
Jardinaud et al. (1995) 또한 옥수수 소포자를 bombardment 기술에 이용하 여 안정적인 형질전환 기술을 확보하였으며, 형질전환된 반수체 로부터 옥수수 재분화체 및
gus (β-glucuronidase) 유전자를 확인 하였다. 한편
Wan et al. (1995)은 약배양으로부터 유래된 캘래 스를 microprojectile bombardment 기술에 적용하였으며, 약배 양에 적합한 유전자형을 선발하여 옥수수 형질전환 효율성을 높이기 위해서 형질전환 연구에 적합한 캘러스를 유도하여 선발 하였다. 일반적으로 약배양은 유전자형에 대한 의존도가 높고 중요하기 때문에 일부 연구에서는 처음부터 androgenic 반응이 매우 높은 유전자형을 선택해서 제초제 저항성 유전자를 지닌 non-androgenic 형질전환체와 교배하여 약배양 형질전환 연구 를 수행하고 있으며, 이 때 non-androgenic 형질전환체는 약배양 을 위하여 공여식물로 사용하였다(
Aulinger et al. 2003). 약과 소포자뿐만 아니라 소포자 유래 액체배양으로부터 원형질체를 분리하여 형질전환 연구에 적용한 결과 유전자가 안정적으로 발현되었고 형질전환 식물체도 확인하였다 (
Shukhapinda et al. 1993). 국내에서 옥수수 약배양 및 소포자를 이용한 형질전환 연구는 매우 드물고 일부 연구자들에 의해서만 진행되고 있으며, 국내 연구 시설에 적합한 약배양 형질전환 시스템 또한 전무한 실정이다. 옥수수 약배양으로부터 형질전환 효율성을 높이기 위해서 다양한 옥수수 품종을 선발 및 선별하는 것도 중요하지만, 연구의 효율을 높이기 위해서는 일반적으로 잘 알려진 androgenic 반응성이 높은 유전자형을 선택하는 것이 더 중요하 다. 이러한 효율성 좋은 품종과의 교배를 통해 옥수수 육종 시스 템을 발달시킨다면 약배양을 이용한 형질전환 연구는 국내 분자 육종 기술 활성화에 기여하고 발전 가능성이 높다고 생각한다.
적 요
옥수수 약배양 기술을 이용하여 DH 계통을 생산하기 위한 전략은 우선 반수체를 생산하여 반수체를 확인한 후, 염색체를 배가시켜 DH 계통을 재현 및 복구하는 것이 약배양의 기본 연구 과정이다. 하지만 현재 우리나라의 옥수수 약배양 기술은 과채류 및 원예작물, 벼, 보리, 밀 등 다른 화본과 작물들에 비하여 연구 업적 및 결과 등이 많지 않으며, 실제로 옥수수 약배양에 적합한 품종 선별 및 선택이 쉽지 않다. 반면 약배양을 통하여 유도된 반수체와 염색체가 배가된 옥수수는 포장에서 수확한 옥수수보다 일반적으로 약하기 때문에 고온 및 저온, 병해충, 제초제 등의 다양한 환경에 대하여 취약한 것이 사실이 다. 하지만 약배양을 이용한 DH 계통 생산은 유용유전자선발이 가능하고 육종 효율성이 높기 때문에, 최종적으로 육성 품종을 신속하게 개발하여 분자육종과 결합하면 연중 육종 시스템을 가동시킬 수 있는 유용한 기내 배양 기술이다. 이러한 옥수수 약배양은 분자육종 및 생명공학에 활용하여 다양하고 우수한 옥수수 신품종을 육성할 수 있는 가능성 높은 기술이며, 국내에 적합한 옥수수 약배양 기술 개발을 위하여 많은 노력이 필요하다.
사 사
본 연구는 농림축산식품부 동남아시아 적응 수출용 옥수수 품종 및 수출 촉진화 기술 개발(213001-04-4-SB920)에 의해 이루어진 것임. 본 논문은 농촌진흥청 공동연구사업(과제번호: PJ00993903)의 지원에 의해 이루어진 것임.
References
- 1. Afele JC, Kannenberg LW. Genetic studies of corn (Zea mays L.) anther culture response. Theor. Appl. Genet 1990. 80: 459-464.
- 2. Ambrus H, Darko E, Szabo L, Bakos F, Király Z, Barnabás B. In vitro microspore selection in maize anther culture with oxidative-stress stimulators. Protoplasma 2006. 228: 87-94.
- 3. Anonymous401 Research Group Primary study on induction of pollen plants of Zea mays. Acta. Genet.Sin 1975. 2: 138-143.
- 4. Antoine-Michard S, Beckert M. Spontaneous versus colchicine-induced chromosome doubling in maize anther culture. Plant Cell, Tissue and Organ Culture 48. 203-207.
- 5. Aulinger IE, Peter SO, Schmid JE, Stamp P. Rapid attainment of a doubled haploid line from transgenic maize (Zea mays L) plants by means of anther culture. In Vitro Cell. Dev. Biol.-Plant 2003. 39: 165-170.
- 6. Barnabás B. In: Maluszynski M, Kasha KJ, Forster BP, Szarejko I. anther culture of maize (Zea mays L). (Eds). Doubled haploid production in crop plants A manual. 2003. Dordrecht, The Netherlands: Kluwer academic publishers, pp. 103-108.
- 7. Barnabás B, Fransz PF, Schel JHN. Ultrastructural studies on pollen embryogenesis in maize (Zea mays L). Plant Cell Rep 1987. 6: 212-215.
- 8. Barnabás B, Obert B, Kovacs G. Colchicine, and efficient genome-doubling agent for maize (Zea mays L) microspores cultured in anthero. Plant Cell Rep 1999. 18: 858-862.
- 9. Beaumont VH, Rocheford TR, Widholm JM. Mapping the anther culture response genes in maize (Zea mays L). Genome 1995. 38: 968-975.
- 10. Büter B, Pescitelli SM, Berger K, Schmid JE, Stamp P. Autoclaved and filter sterilized liquid media in maize anther culture significance of activated charcoal. Plant Cell Rep 1993. 13: 79-82.
- 11. Büter B, Schmid JE, Stamp P. Effects of L-proline and post-plating temperature treatment on maize (Zea mays L) anther culture. Plant Cell Rep 1991. 10: 325-328.
- 12. Chang MT, Coe EH. Kriz AL, Larkins A. Doubled haploids. Biotechnology in Agriculture and Forestry. Molecular Genetic Approaches to Maize Improvement 2009. 63. Berlin Heidelberg, Springer Verlag. 127-142.
- 13. Chase SS. Monoploids and monoploid-derivatives in maize (Zea mays L). The Botanical Review 1969. 35: 117-167.
- 14. Chu CC, Wang CC, Hsu CS, Yin KC, Chu YC, Bi FY. Establishment of an efficient medium for anther culture of rice through comparative experiments on nitrogen sources. Sci. Sin 1975. 18: 484-490.
- 15. Coumans MP, Sohota S, Swanson EB. Plant development from isolated microspores of Zea mays L. Plant Cell Rep 1989. 7: 618-621.
- 16. Das UR, Hadiuzzaman S, Sarker RH. Effects of microspore stage, pre- and post temperature and donor’s environment on maize (Zea mays L) anther culture response. Plant Tissue Cult 2002. 12(1):37-47.
- 17. Dieu E, Beckert M. Further studies of androgenetic embryo production and plant regeneration from in vitro cultured anthers in maize (Zea mays L). Maydica 1986. 31: 245-259.
- 18. Dhekney SA, Li ZT, Compton ME, Gray DJ. Optimizing initiation and maintenance of vitis embryogenic cultures. Hort. Science 2009. 44(5):1400-1406.
- 19. Farnham MW. Doubled-haploid broccoli production using anther culture effect of anther source and seed set characteristics of derived lines. J. Amer. Soc. Hort. Sci 1998. 123: 73-77.
- 20. Feng XR, Wolyn DJ. High frequency production of haploid embryos in asparagus anther culture. Plant Cell Rep 1991. 10: 574-578.
- 21. Gaillard A, Matthys-Rochon E, Dumas C. Selection of microspore derived embryogenic structures in maize related to transformation potential by microinjection. Botanica Acta 1992. 105: 313-318.
- 22. Gaillard A, Vergne P, Beckert M. Optimization of maize microspore isolation and culture conditions for reliable plant regeneration. Plant Cell Rep 1991. 10: 55-58.
- 23. Geiger HH, Gordillo GA. Doubled haploids in hybrid maize breeding. Maydica 2009. 54: 485-499.
- 24. Genovesi AD. Bajaj YPS. Maize (Zea mays L.): in vitro production of haploids. Biotechnology in Agriculture and Forestry. Haploids in crop improvement I 1990. 12. Berlin, Springer. 176-203.
- 25. Genovesi AD, Collins GB. In vitro production of haploid plants of corn via anther culture. Crop Sci 1982. 22: 1137-1144.
- 26. Germanà MA. Gametic embryogenesis and haploid technology as valuable support to plant breeding. Plant Cell Rep 2011. 30: 839-857.
- 27. Gu HH, Hagberg P, Zhou WJ. Cold treatment enhances microspore embryogenesis oilseed rapa (Brassica napus L.). Plant Growth Regul 2004. 42: 137-143.
- 28. Guha S, Maheshwari SC. In vitro production of embryos from anthers of Datura in vitro. Nature 1964. 204: 497.
- 29. Guha S, Maheshwari SC. Cell division and differentiation of embryos in the pollen grains of Datura in vitro. Nature 1966. 212: 97-98.
- 30. Häntzschel KR, Weber G. Blockage of mitosis in maize root tips using colchicines-alternatives. Protoplasma 2010. 241: 99-104.
- 31. Hassan L, Ahmad SD, Okimus A. The direct regeneration of maize haploids through anther culture. Journal of Biological Sciences 2001. 1(10):900-901.
- 32. Hosseini SZ, Ismaili A, Mohammadi PP. Improvement of maize (Zea mays L) anther culture embryogenesis and direct regeneration by different plant growth regulators and gelling agent. Journal of Applied Biotechnology Reports 2014. 1(1):17-21.
- 33. Hosseini SZ, Mohammadi PP, Cheshmeali GR. Effect of induction medium on embryogenesis of maize anther culture and increase of normal haploid plantlet regeneration with wathman filter paper. Iranian Journal of Plant Physiology 2015. 5(4):1505-1512.
- 34. Indrianto A, Barinova I, Touraev A, Heberle-Bors E. Tracking individual wheat microspores in vitro identification of embryogeneic microspores and body axis formation in the embryo. Planta 2001. 212: 163-174.
- 35. Islam SMS. Evaluation of regeneration potential of microspore derived embryoids in wheat (Triticum aestivum L.). Plant Omics. J 2010. 3: 149-153.
- 36. Islam SMS. The effect of colchicine pretreatment on isolated microspore culture of wheat (Triticum aestivum L.). Aus. J. Crop Sci 2010. 4: 660-665.
- 37. Ismaili A, Mohammadi PP. Effect of genotype, induction medium, carbohydrate source, and polyethylene glycol on embryogenesis in maize (Zea mays L) anther culture. Acta Physiol. Plant 2016. 38(3):1-8.
- 38. Jardinaud MF, Souvé Alibert G, Beckert M. uidA gene transfer and expression in maize microspores using the biolistic method. Protoplasma 1995. 187: 138-143.
- 39. Jo MH, Ham IK, Park MY, Kim TI, Lim YP, Lee EM. Seed production ability of doubled haploid plants through microspore culture in Chinese cabbage (Brassica rapa, L. ssp. pekinensis). Korean. J. Hort. Sci. Technol 2012. 30(5):573-578.
- 40. Jumpatong C, Boonyai P, Sangduen N, Thriaporn R, Saisingtong S, Büter B. anther culture–a new tool for the generation of double haploid, homozygous maize in Thailand. Thai. J. Agric. Sci 1996. 29: 469-487.
- 41. Kahrizi D, Moieni A, Bozorgipour R. Effect of genotype and carbohydrate source on androgenesis of hexaploid wheat (Triticum aestivum L). Seed Plant 2000. 16: 41-51.
- 42. Karsai I, Bedo Z. Relationship between anther culture response and aluminium tolerance in wheat (Triticum aestivum L). Euphytica 1998. 100: 249-252.
- 43. Kasha KJ, Ziauddin A, Cho UH. In: Gustafson JP Haploids in cereal improvement anther and microspore culture. (Ed). In gene manipulation in plant improvement II. 1990. New York: Plenum Press, pp. 213-235.
- 44. Khavari KS, Moieni A, Mousavi A, Golbashy M. Study on embryogenesis response of maize (Zea mays L) genotypes to anther culture. Agricultural Biotechnology 2011. 10(1):35-46.
- 45. Ku M, Cheng W, Kuo L, Kuan Y, An H, Huang C. Studies on induction of pollen plants in maize. Proc. Symp. anther Cult 1997. Beijing, China, Scientific Press. 18-26.
- 46. Kuo CS, Sun AC, Wang YY, Gui YL, Gu SR, Miao SH. Studies on induction of pollen plants and androgenesis in maize. Acta. Bot. Sin 1978. 20(3):204-209.
- 47. Marchic A, Antoine-Michard S, Bordes J, Pollacsek M, Murigneux A, Beckert M. Genetic improvement of antheranther culture response in maize relationship with molecular, Mendelian and agronomic traits. Theoretical and Applied Genetics 1998. 97: 520-525.
- 48. Mohammadi PP, Moieni A, Javaran MJ. Colchicine induced embryogenesis and doubled haploid production in maize (Zea mays L) anther culture. Iranian. J. Biotech 2007. 5(3):140-146.
- 49. Nitsch C. Reinert J, Bajaj YPS. Culture of isolated microspores. Applied and fundamental aspects of plant cell, tissue, and organ culture 1977. Berlin Heidelberg New York, Springer Verlag. 66-91.
- 50. Nitsch C, Andersen S, Godard M, Neuffer MG, Sheridan WF. In: Earle ED, Demarly Y. Production of haploid plants of Zea mays and Pennisetum through androgenesis. (Eds). Variability in plants regenerated from tissue culture. 1982. New York: Praeger Press, pp. 69-91.
- 51. Obert B, Barnabás B. Colchicine induced embryogenesis in maize. Plant Cell Tiss. Organ. Cult 2004. 77: 283-285.
- 52. Pechan PM, Smykal P. Androgenesis Affecting the fate of the male gametophyte. Physiologia Plantarum 2001. 111: 1-8.
- 53. Pescitelli SM, Johnson CD, Petolino JF. Isolated microspore culture of maize effects of isolation technique, reduced temperature, and sucrose level. Plant Cell Rep 1990. 8: 628-631.
- 54. Petolino JF, Jones AM. anther culture of elite genotypes of maize. Crop Science 1986. 26: 1072-1074.
- 55. Petolino JF, Jones AM, Thompson SA. Selection for increased anther culture response in maize. Theor. Appl. Genet 1988. 76: 157-159.
- 56. Petolino JF, Thompson SA. Genetic analysis of anther culture response in maize. Theor. Appl. Genet 1987. 74: 284-286.
- 57. Portemer V, Renne C, Guillebaux A, Mercier R. Large genetic screens for gynogenesis and androgenesis haploid inducers in Arabidopsis thaliana failed to identify mutants. Front. Plant Sci 2015. 6: 147.
- 58. Prasanna BM, Chaikam V, Mahuku G. Doubled haploid technology in maize breeding Theory and practice. CIMMYT, Mexico, D.F 2012.
- 59. Rober FK, Gordillo GA, Geiger HH. In vivo haploid induction in maize-performance of new inducers and significance of doubled haploid lines in hybrid breeding. Maydica 2005. 50: 275-283.
- 60. Saisingtong S, Schmid JE, Stamp P, Buter B. Colchicine-mediated chromosome doubling during anther culture of maize (Zea mays L.). Theor. Appl. Genet 1996. 92: 1017-1023.
- 61. Seo MS, Sohn SH, Park BS, Ko HC, Jin M. Efficiency of microspore embryogenesis in Brassica rapa using different genotypes and culture conditions. J. Plant Biotechnol 2014. 41: 116-122.
- 62. Shukhapinda K, Kozuch ME, Rubin-Wilson B, Ainley WM, Merlo DJ. Transformation pf maize (Zea mays L) protoplasts and regeneration of haploid transgenic plants. Plant Cell Rep 1993. 13: 63-68.
- 63. Spitko T, Sagi L, Pintér J, Marton LC, Barnabás B. Haploid regeneration aptitude of maize (Zea mays L) lines of various origin and of their hybrids. Maydica 2006. 51: 537-542.
- 64. Torp AM, Andersen SB. In: Touraev A, Forster BP, Jain SM. Albinism in microspore culture. (Eds). Advances in haploid production in higher plants. 2009. The Netherlands: Springer, pp. 155-160.
- 65. Touraev A, Ilham A, Vicente O, Heberle-Bors E. Stress-induced microspore embryogenesis in tobacco an optimized system for molecular studies. Plant Cell Rep 1996. 15: 561-565.
- 66. Tulecke W. A haploid tissue culture from the female gametophyte of Ginkgo biloba. Nature 1964. 203: 94-95.
- 67. Uvácková L, Takác T, Boehm N, Obert B, Šamaj J. Proteomic and biochemical analysis of maize anthers after cold pretreatment and induction of androgenesis reveals and important role of anti-oxidative enzymes. Journal of Proteomics 2012. 75: 1886-1894.
- 68. Wan Y, Duncan DR, Rayburn AL, Petolino JF, Widholm JM. The use of antimicrotubule herbicides for the production of doubled haploid plants from anther-derived maize callus. Theor. Appl. Gent 1991. 81: 205-211.
- 69. Wan Y, Petolino JF, Widholm JM. Efficient production of doubled haploid plants through colchicine treatment of anther-derived maize callus. Theor. Appl. Genet 1989. 77: 889-892.
- 70. Wan Y, Widholm JM, Lemaux PG. Type I callus as a bombardment target for generating fertile transgenic maize (Zea mays L.). Planta 1995. 196: 7-14.