적요
Soybean [Glycine max. (L.) Merr] is one of the most important legumes in the world. However, soybean varieties are sensitive to flooding stress and their seed yields are substantially reduced in response to the flooding stress. 192 soybean germplasm collection was screened to identify flooding tolerant germplasm at an early vegetative growth stage (V1). Soybean plants at V1 stage were waterlogged for 4 to 10 days. To evaluate flooding tolerance, survival rate were investigated as a time dependent manner. Jangbaegkong, Danbaegkong, Sowonkongkong, Socheong2 and Suwon269 showed flooding tolerance, while Shillog, T201, T181, NTS1116 and HP-963 showed flooding sensitivity. We also investigated effects of flooding stress on soybean morphology. The adventitious root development was greatly increased in flooding tolerant plants compared to it in flooding sensitive plants. In addition, root length and root number were analyzed. The significant reduction of root length and root number was observed in flooding sensitive plants. Thus, these results indicate that the morphological changes in roots are important for acclimation to flooding stress. Taken together, the relationship between the morphological changes in the roots and flooding tolerance may be useful in selecting a flooding tolerant soybean germplasm.
서 언
콩은 농업적 측면에서 지력유지 및 증진에 유용한 작물이 며, 쌀과 더불어 식량자원으로서 우리나라 농업의 중요한 부 분을 차지하고 있다. 그러나 1960년대 이후부터 콩 재배면적 은 감소하기 시작하여, 2000년대 이후에는 콩 자급률이 6% 대에 불과한 실정이다. 우리나라에서 여름철(6월~8월) 강수 량은 연 강수량의 40~60%를 차지하며, 여름철 집중 호우로 인한 과습 피해가 빈번히 발생하고 있다. 특히 평야지대에서 배수 불량으로 인한 과습 피해는 콩의 생육 저하와 수량 감소 의 주원인이 되고 있다.
과습에 의한 피해는 주로 뿌리에서 나타난다. 그 결과, 식 물체의 지상부는 시듦 현상을 보이는데, 이러한 현상은 산소 결핍에 의한 독성대사물질의 생성 및 양분결핍 등에 기인하 는 것으로 알려져 있다(Akiko et al. 2009,
Armstrong et al. 1994). 과습 상태가 지속되면 여러 가지 피해 현상이 나타난 다. 1) 과습으로 인해 뿌리의 생육이 나빠지며, 질소고정 능력 이 낮아진다(
Fehr et al. 1971). 즉, 과습 상태에서는 산소가 부족하게 되어 뿌리는 지표면에 분포하게 되며, 길이는 짧고 연약해지며, 껍질부분은 굳어지게 된다. 따라서 양분과 수분 의 흡수 장해로 인해 뿌리혹의 무게와 근류균의 활성이 떨어 지며, 엽록소의 함량, 지상부와 지하부의 생체중 등이 감소한 다. 2) 과습으로 인해 식물체의 양분흡수와 물질이동이 감소 한다(
Bacanamwo and Purcell, 1999). 과습에 의한 양분흡수 는 생육시기, 양분요소, 피해 부위에 따라 다르게 나타나지만 전체적인 질소, 석회, 마그네슘, 칼리의 흡수는 저해된다. 3) 과습으로 인해 생육이 저해되고 수량성이 감소한다(
Scott et al. 1989). 과습으로 인해 잎의 엽록소 함량이 감소하며, 아래 잎부터 누렇게 변하기 시작하여 상위 잎으로 진전되거나 위 조 현상이 발생하게 된다(
Lee et al. 2010). 콩의 경우, 과습 기간이 5일 이상 일 때, 총엽록소 함량, 활성근류의 양, 근류 의 질소고정능력 등이 급격히 감소하며, 7일 이상일 때, 모든 생리기능이 현저히 저하되어 수량이 약 5% 정도 감소되며, 14일 이상일 때, 수량이 7~10% 감소한다. 또한, 이러한 과 습 피해는 영양생장기보다 개화기에 더욱 심각하다고 알려져 있다(
Kim et al. 2010). 식물체가 습해를 받게 되면, 한발을 받을 때와 마찬가지로 엽수분 potential이 감소하게 된다. 따 라서 수분손실을 최소화하기 위해 기공을 닫게 되어 기공 전 도도가 감소한다. 이로 인해 가스와 수분의 출입이 원활히 이 루어지지 못하게 되어 광합성속도, 증산량 및 수분이용효율도 감소한다(
Bennet & Albrecht, 1984;
Boyer, 1970).
일반적으로 식물체가 과습을 받게 되면, 전체적인 성장이 저해 된다. 그러나 특정 조직의 발달등과 같은 형태적인 변화 를 통해 과습 환경에 적응하게 된다. 그 예로 부정근의 발달 은 과습에 대한 식물체의 초기 적응의 한 현상이다. 식물체가 침수 피해를 입게 되면, 산소분압이 비교적 높은 지표면 가까 이에서 부정근의 발달을 증가시킨다. 과습에 강한 식물체일수 록 부정근의 수가 많아지며, 생육후기의 양분과 수분 흡수 능 력이 증대된다(
Thomas et al. 2005)과습에 의한 부정근 발달 은 ethylene에 의해 촉진되며, 고농도의 ethylene은 부정근이 형성되는 조직 생체 내에서 유래한 auxin의 감응도를 높여준 다(
Visser et al. 1996). 단기간의 과습인 경우 뿌리가 짧고, 굵고, 약해지며 cortical cell 조직이 목화(lignification)되거나 목전화(suberization)되기 때문에 양분과 수분의 흡수가 저해 된다(
Lee et al. 2010).
우리나라 기상여건상 콩 생육초기인 6~8월에 습해로 인 한 입모율 및 생육 저하 등의 피해가 빈번하게 발생하고 있다. 또한 최근 지구온난화에 따른 기상재해 빈도 및 강도가 갈수 록 심화되고 있다. 그러나 콩 습해에 대한 연구는 매우 빈약 한 실정이다. 따라서 본 시험은 습해 저항성 유전자원을 선발 하고, 향후 내습성 콩 품종 조기육성을 위한 기초자료를 제공 하고자 수행하였다.
재료 및 방법
실험재료
본 실험에서는 국립식량과학원 기능성작물부 대구시험지 에서 2011년 6월 중순에 파종하여 수확된 콩 주요 교배 모부 본 및 보유 유전자원 등 192점으로 습해 저항성 자원을 2012 년에 1차 선발하였다. 1차로 선발된 습해 저항성 및 감수성 자원 각각 5점에 대해 2013년에 재현성 및 표현형 조사를 하 였다.
습해 처리 및 뿌리 생육조사
본 실험은 국립식량과학원 기능성작물부 대구시험지에 설 치된 가림막 온실에서 2012년과 2013년에 수행하였다. 가림 막 온실은 외부와 동일한 조건을 위해 네 측면을 개방하였으 며, 천정에 비닐을 설치하여 자연 강우를 배제하였다. 가로 1 m, 세로 2 m, 높이 1 m의 bed에 배수를 위해 자갈을 30 cm 정 도로 채우고, 60 cm 정도로 밭흙을 채웠다. 각 자원당 1주 3 본으로 파종하였으며, 발아 후 1개체를 남기고 솎아주었다. 5 개체 반복으로 습해 처리구 3반복과 대조구 1반복으로 각각 의 bed에 배치하였다. 과습처리는 V1~V2 stage에 10일 이 상, 지표 위 5~10 cm로 침수 처리하여 과습을 유발하였다. 대조구는 정상적으로 관수를 하였다. 습해 저항성 및 감수성 은 침수처리에 의한 생존율로 구분하였다. 또한, 습해 처리에 의한 저항성과 감수성의 뿌리 생육을 관찰하기 위해 과습 처 리 종료 약 3주 후 각 자원당 3개체씩을 채취하여 부정근 수, 뿌리 길이, 뿌리 수를 조사하였다.
RT-PCR과 qRT-PCR에 의한 습해관련 유전자 발현 분석
RT-PCR과 qRT-PCR에 의한 유전자 발현분석을 위한 primer 설계는 Primer 3 (
http://frodo.wi.mit.edu/, version 0.4) 프 로그램을 사용하였다. PCR 증폭산물의 크기는 300~500 bp, Tm값과 GC%, PCR에서 mismatch 확률, primer dimer 생성 유무 등을 검토하여 RT-PCR 에 적합한 primer쌍들을 설계 하였다. 습해관련 유전자에 대한 primer들은
APX1 (forward primer, GAGCGCTGATTACCAGAAGG, reverse primer, CTTTGAGTGGCTCCAAAAGC),
APX2 (forward primer, AACTTACGACGTGAGCTCGAAG, reverse primer, GAG CCTCAGCGTAATCAGCGAAG)와
Adh (forward primer, AT GTCGAGCACAGCTGGCCA, reverse primer, CTGTGCA AATTCCACAAC)으로 작성하였다(
Kausar et al. 2012,
Shi et al. 2008,
Komatsu et al. 2011). 또한 internal control로 사용된
GmACT11 (forward primer, CTTCAGGCATTCAC GAGACA, reverse primer, GTGCACAATTGATGGACCAG) 을 제작하여 사용하였다. 습해 처리 전(0일)과 습해 처리 후 3, 5, 9일로 잎 시료를 채취하여 total RNA를 추출하여 cDNA 를 합성하였다. 1 μg total RNA, 50 μM oligo (dT) primer 를 혼합하여 70°C에서 5분간 가열한 후 바로 ice에 정치하였다. 0.5 mM dNTP mix, 5x PrimeScript buffer, 20 units RNase inhibitor (Promega, USA), 100 units PrimeScript reverse transcriptase (Takara, Japan)을 이용하여 총 반응용액 20 μl 를 가지고 42°C에서 60분간 역전사 반응을 수행하였다. cDNA 합성 후 반응액을 70°C에서 15분간 정치한 다음 RT-PCR에 사용하였다. RT-PCR 반응조건은 96°C 5분, 96°C 30초, 60°C 30초, 72°C 45초씩 30cycle 반응시킨 후, 72°C에서 10분간 반응하여 종결시켰다(
Koo et al. 2009).
GmACT11을 internal control로 이용하여 다른 유전자와의 양적 발현을 분석하였으 며, 각각 1/10 volume의 반응산물을 2% agarose gel에 전기 영동하여 발현 정도를 확인하였다. qRT-PCR는 위의 각 primer 와 SYBR Green SuperMix (Bio-Rad Lab, Hercules, CA, USA)을 사용하여 7300 Real Time PCR system (Applied Biosystems, Foster City, CA, USA)으로 유전자 발현의 정 량분석을 실시하였다. 습해 관련 유전자의 계산된 threshold cycle (Ct)는 대조 유전자인
ACT11의 Ct에 의해 표준화하였 으며, 각 유전자들의 상대적인 값은 ΔΔCt 방법을 이용하여 계산하였다(
Livak and Schmittgen, 2001).
모든 데이터는 3회 반복 측정하였으며, mean±SD로 표현 하였다. 통계분석은 R version 3.0.2 program을 이용하여, 각 처리군의 평균과 표준편차를 산출하고 Duncan's multiple range test를 이용하여 유의성을 검정하였다.
결과 및 고찰
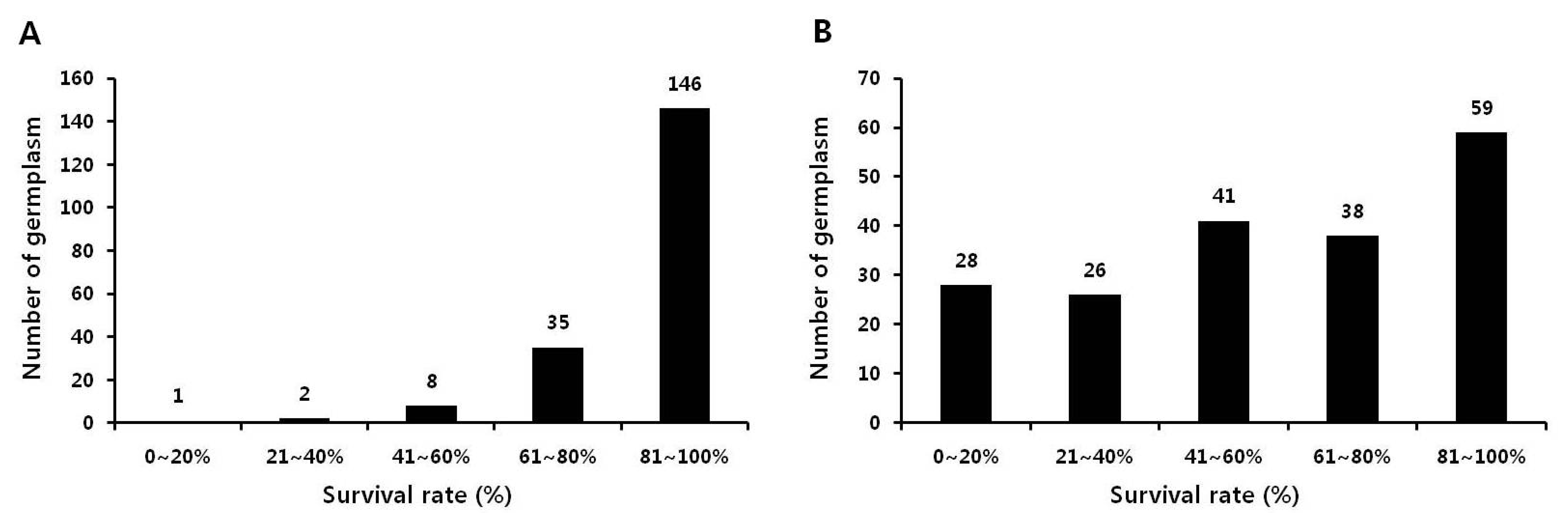
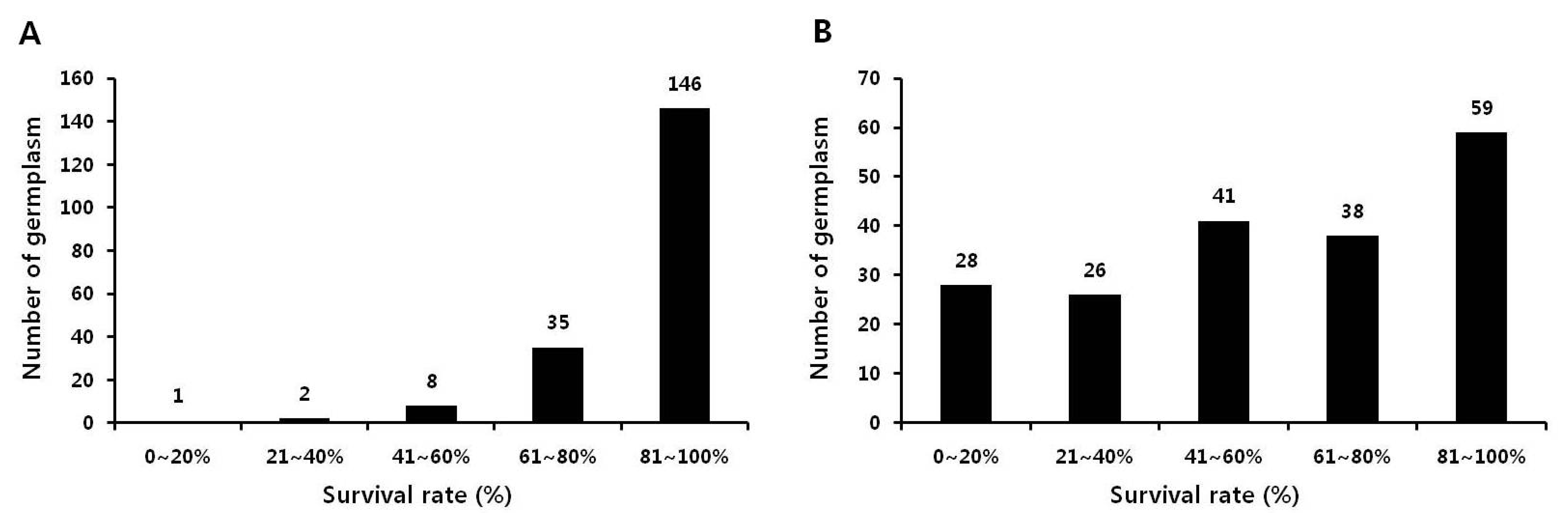
본 실험에서는 국립식량과학원 기능성작물부 대구시험지 에서 국내외 품종 및 유전자원 등 192종을 대상으로 2012년 에 습해 저항성 자원을 1차 선발하였다. 습해 저항성 자원 선 발을 위해 습해 저항성과 감수성으로 보고된 PI408105A와 S99-2281을 각각 대조구로 사용하였다(Lakshmi 등 2010). 먼저, 상토와 밭토양에서 키운 생육초기(V1 stage) 식물을 각 각 침수 처리하였다. 밭토양에 비해 상토에서 키운 식물은 침 수처리에 적합하지 않음을 확인하였다. 따라서 본 실험에서는 밭토양에 키운 식물을 이용하여 실험을 진행하였다. 192종의 생육초기 식물에 4, 5, 6, 7, 10일간 침수처리 후 생존율을 조 사하였다. 생존율은 전체 개체 중 생존한 개체를 백분율로 계 산하였다. 4일간 침수 처리했을 때에는 146여점의 생존율이 80% 이상으로 습해에 대한 피해가 거의 발생하지 않았다 (Fig.
1A). 하지만 침수 기간이 길어질수록 생존율이 낮아지 는 경향을 보였다. 특히, 10일간 침수 처리시 95여점의 유전 자원이 60% 이하의 생존율을 나타내었다(Fig.
1B). 이 결과 를 통해, 생육초기에 습해 저항성과 감수성 자원을 선발하기 위해서는 10일 이상 침수 처리하는 것이 적당할 것으로 사료 된다. 침수처리 후 생존율 조사 결과를 토대로, 각 침수 기간 에서 생존율이 80% 이상인 자원들을 습해 저항성 자원으로 선발하였고, 생존율이 40% 이하인 자원들을 습해 감수성 자 원으로 선발하였다(Fig.
1). 장백콩(Jangbaegkong), 단백콩 (Danbaegkong), 소원콩(Sowonkong), 소청2호(Socheong2), 수원269(Suwon269) 등을 습해 저항성 자원으로, 신록(Shillog), T181, T201, NTS1116, HP-963 등을 습해 감수성 자원으로 각각 선발하였다. 습해 저항성과 감수성으로 보고된 PI408105A 와 S99-2281의 생존율은 각각 63%와 55%로 습해에 대한 중 도 저항성으로 분류되었으며, 이는 선발 모지와 우리나라의 재배환경이 다르기 때문으로 사료된다.
Fig. 1.Survival rate of soybean seedling under flooding stress for 4 days (A) and 10 days (B). Soybean plants at V1 stage were waterlogged.
1차 선발된 자원들을 대상으로 습해처리에 대한 재현성 검 정을 실시하였다. 그 결과, 습해 저항성 및 감수성 자원들의 생존율은 1차 실험과 유사한 결과를 보였다. 대조구에 비해 처리구에서 저항성과 감수성 자원 모두 지상부가 왜소해지는 현상을 나타내었다. 하지만 감수성 자원에서는 저항성 자원에 비해 황화현상과 위조현상이 심하게 발생하였다. 저항성 자원 에서 약하게 황화현상이 발생하였지만 배수 후 잎색이 회복 되는 현상을 확인할 수 있었다(Fig.
2A). 선발된 습해 저항성 자원들은 71% 이상의 생존율을 나타내었으며, 특히 장백콩 과 소청2호는 100%의 생존율을 보였다. 이에 반해 습해 감수 성 자원들은 25% 이하의 생존율을 나타내었다(Fig.
2B).
Fig. 2.Phenotype (A) and survival rate (B) by flooding stress. A, Phenotype of tolerance and sensitivity for flooding stress. In each panel left plant is a control and right plant is treated by flooding. B, Survival rate of tolerance and sensitivity for flooding stress.

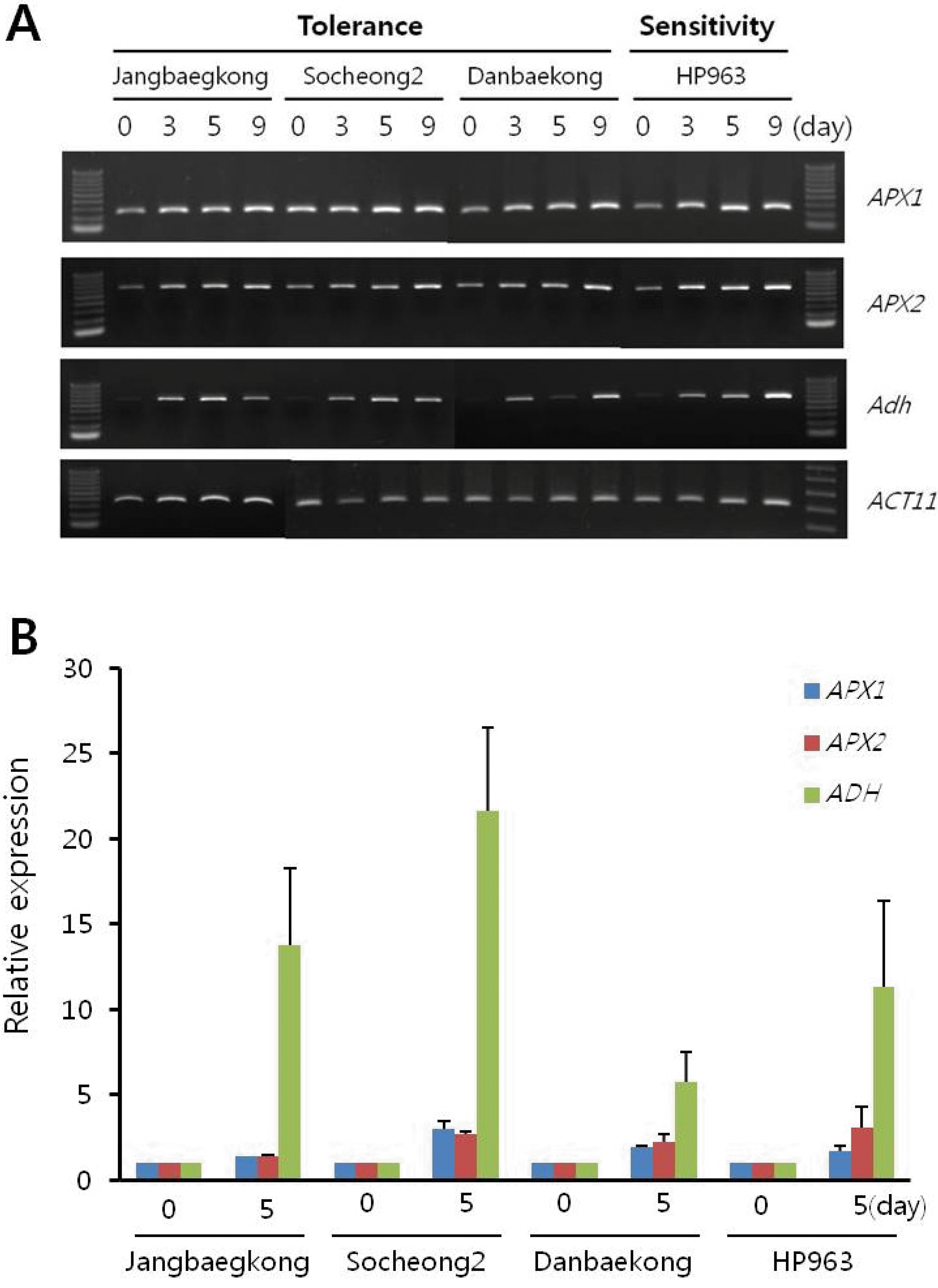
이러한 현상이 습해에 의해 유도되는 현상인지 알아보기 위해 습해 처리에 의해 발현이 증가된다고 알려진
APX1 (
ASCORBATE PEROXIDASE 1),
APX2 (
ASCORBATE PEROXIDASE 2),
Adh (
Alcohol dehydrogenase) 등의 유전 자들에 대해 발현 변화를 조사하였다(
Kausar et al. 2012;
Shi et al. 2008;
Komatsu et al. 2011). Fig.
3에서 보이는 바와 같이
APX1, APX2, Adh의 발현은 습해 처리 전에 비해 습해 처리 후에 발현이 증가 되는 것을 확인 할 수 있었다. 이를 통 해 습해 처리가 정상적으로 처리되었음을 알 수 있다.
Fig. 3.Expression analysis of APX1, APX2, and ADH in floodingtolerant and -sensitive germplasms during flooding. A, RT-PCR analysis by flooding treatment for 0, 3, 5, or 9 day. ACT11 gene was used as an internal control in the RT-PCR. B, qRT-PCR analysis by flooding treatment for 0 and 5 day. Relative expression measured by real-time qRT-PCR is normalized to ACT11. Error bars indicate s.e.m.
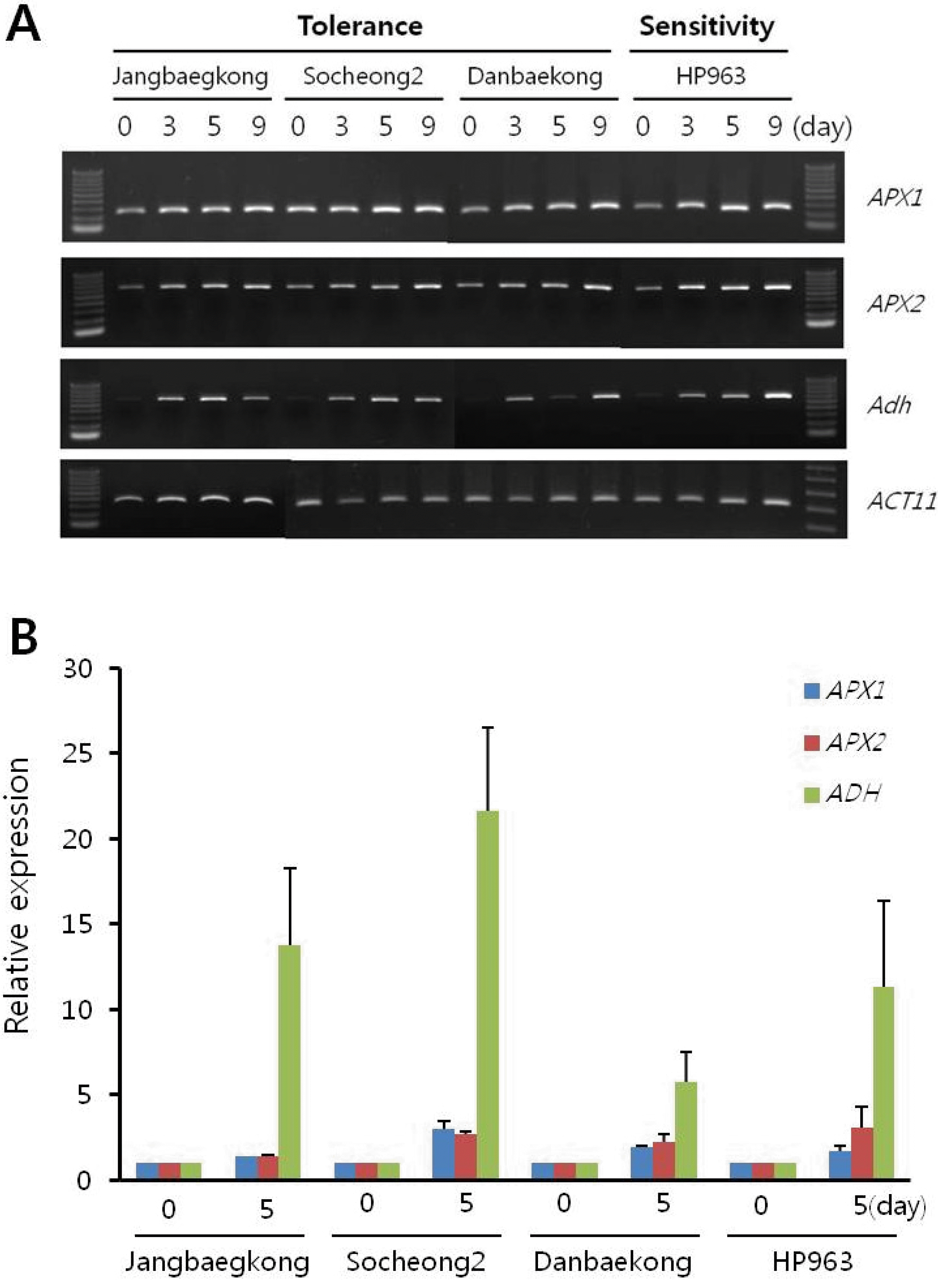
또한 습해에 의한 식물체의 표현형 변화가 식물체의 침수 되는 정도 차에 의해 나타나는 현상인지를 알아보기 위해 배 축 높이를 측정하였다. Fig.
4A에서 보이는 결과와 같이, 습 해 처리 후 나타나는 표현형은 침수되는 높이와는 상관이 없 음을 확인하였다. 따라서 습해 처리 후 나타나는 표현형은 각 자원의 습해에 대한 유전적 특성임을 알 수 있다.
Fig. 4.Hypocotyl Length and root morphology by flooding stress. A, Hypocotyl length was measured below the cotyledons and above the radicle. B, Adventitious roots were counted in hypocotyl. C and D, Root number and length are compared to plants of control and flooding treatment. Square and triangle indicate the control and flooding plant, respectively. Error bars represent standard error. Any means in the same column followed by the same letter are not significantly (p<0.05) different by Duncan's multiple range test.
최근의 연구 결과에 의하면, 습해에 의해 부정근이 발달된 다고 보고되어 있다(
Thomas et al. 2005). 이를 근거로 선발 된 습해 저항성 및 감수성 자원에 대해 습해 처리 후 부정근 의 발달 정도를 조사하였다(Fig.
4B). 습해 저항성 자원에서 부정근 수는 17.3~41.3개로 조사되었다. 특히 장백콩은 습해 처리 후 평균적으로 41.3개로 가장 왕성한 부정근의 발달을 보였다. 이에 반해 습해 감수성 자원에서 부정근 수는 0~9개 로 조사되었다. 이 결과는 습해 감수성 자원에 비해 습해 저항 성 자원에서 부정근의 발달이 촉진되고 있음을 보여주고 있다. 따라서 본 실험의 결과는 침수된 식물체에서의 부정근 발달은 습해에 대한 검정지표로 사용될 수 있음을 시사해주고 있다.
식물체가 습해를 받게 되면 뿌리의 생육과 활력이 낮아진 다. 습해처리 후 뿌리의 생육을 확인하기 위해 습해 저항성 및 감수성 자원에서 측근과 주근의 발달을 조사하였다(Fig
3C and
3D). 습해를 처리하지 않은 대조구에서 감수성 자원 인 T201과 신록의 측근 수는 상대적으로 높았지만, 이 외의 저항성 및 감수성 자원의 측근 수는 비슷하게 조사되었다. 습 해를 처리했을 때 감수성 자원의 측근 수는 대조구에 비해 37 ~65%로 감소하였고, 저항성 자원의 측근 수는 8~67% 증 가함을 보였다(Fig.
3C). 이는 습해 감수성 자원에서는 습해 처리에 의해 측근의 발육이 저해되지만, 습해 저항성 자원에 서는 생존을 위한 산소와 영양분 흡수를 위해 측근 발달이 더 욱 유도 되어지는 것으로 사료된다. 주근 길이는 대조구에서 평균 27.6~40.7 cm로 감수성과 저항성 자원 간에 차이가 거 의 없었다. 하지만 습해를 처리하였을 때, 습해 저항성 자원의 주근 길이는 대조구에 비해 0~25% 정도 줄어든 반면 감수 성 자원의 주근 길이는 44~60% 정도 줄어든 것으로 조사되 었다(Fig.
3D). 이는 습해 감수성 자원이 저항성 자원에 비해 습해에 의해 뿌리 생육이 저해됨을 알 수 있다. 이 결과로 미 루어 뿌리의 생육 상태가 습해 검정지표로 사용될 수 있음을 시사해주고 있다.
현재까지 콩에서 습해에 의한 분자생물학적 조절 기작은 거의 연구가 되어있지 않은 실정이다. 따라서 습해에 대한 분 자생물학적 조절 기작의 구명은 습해 저항성 품종 육성에 큰 도움이 될 것이라 여겨진다.
적 요
본 연구는 우리나라 기상여건을 고려하여 콩 생육초기에 습해 저항성 자원을 선발하여 내습성 콩 품종의 조기육성 및 유전적 요인을 구명하기 위한 기초자료를 제공하고자 수행하 였던 바, 그 결과를 요약하면 다음과 같다.
국내 콩 주요 품종 및 보존 유전자원 등 192점에 대한 담 수처리에 의한 생존율에 따라 습해 저항성과 감수성으로 분류하였다. 그 결과, 장백콩, 단백콩, 소원콩, 소청2호, 수 원269는 습해 저항성 자원으로 선발되었다. 신록, T181, T201, NTS1116, HP-963은 습해 감수성 자원으로 선발되 었다.
습해처리에 의해 발현이 증가된다고 보고된 유전자들 APX1, APX2, Adh의 발현을 분석한 결과 유전자들의 발현이 증 가하였으며, 이로 미루어 습해처리가 정상적으로 처리되었 음을 알 수 있다.
1차 선발된 습해 저항성 및 감수성 자원에 습해를 처리하 였을 때, 저항성 자원은 생존율이 71% 이상, 감수성 자원 은 25% 이하의 생존율을 나타내었다. 1차 선발된 습해 저 항성 및 감수성 자원에 대한 재현성을 확인하였다.
부정근은 습해 저항성 자원에서 17.3~41.3개, 감수성 자 원에서 0~9개를 나타내었다. 습해 감수성 자원에 비해 습 해 저항성 자원에서 부정근이 상대적으로 많이 발달함을 확인할 수 있었다.
주근의 길이는 저항성 자원에서는 0~25% 정도 감소한 반면 감수성 자원에서는 44~60% 감소한 것으로 조사되 었다. 측근의 수는 감수성 자원에서는 37~65%로 감소하 였지만 저항성 자원에서는 8~67% 증가하였다.
사 사
본 논문은 농촌진흥청 연구사업(세부과제명: 콩 내습성 유 전자원 선발 및 분자표지 개발, 세부과제번호: PJ009291012014) 의 지원에 의해 이루어진 것임.
References
- 1. Armstrong W, Brändle B, Jackson MB. Mechanisms of flood tolerance in plants. Acta Bot. Néerl 1994. 43: 307-58.
- 2. Bacanamwo M, Purcell LC. Soybean dry matter and N accumulation responses to flooding stress. N sources and hypoxia. J. Exp. Bot 1999. 50: 689-696.
- 3. Bennett JM, Albrecht SL. Drought and flooding effects on N2 fixation, water relations and diffusive resistance of soybean. Agron. J 1984. 76: 735-740.
- 4. Boyer JS. Differing sensitivity of photosynthesis to low leaf water potentials in corn and soybean. Plant Physiol 1970. 46: 234-239.
- 5. Fehr WR, Caviness CE, Burmood DT, Pennington JS. Stages of development descriptions for soybeans, Glycine max (L.). Merrill. Crop Sci 1971. 11: 929-930.
- 6. Hashiguchi A, Sakata K, Komatsu S. Proteome analysis of early-stage soybean seedlings under flooding stress. J. Proteome Res 2009. 8: 2058-2069.
- 7. Kausar R, Hossain Z, Makino T, Komatsu S. Characterization of ascorbate peroxidase in soybean under flooding and drought stresses. Mol. Biol. Rep 2012. 39: 10573-10579.
- 8. Komatsu S, Thibaut D, Hiraga S, Kato M, Chiba M, Hashiguchi A, Tougou M, Shimamura S, Yasue H. Characterization of a novel flooding stress-responsive alcohol dehydrogenase expressed in soybean roots. Plant Mol. Biol 2011. 77: 309-322.
- 9. Koo SC, Choi MS, Chun HJ, Shin DB, Park BS, Kim YH, Park HM, Seo HS, Song JT, Kang KY, Yun DJ, Chung WS, Cho MJ, Kim MC. The calmodulin- binding transcription factor OsCBT suppresses defense responses to pathogens in rice. Mol. Cells 2009. 27: 563-570.
- 10. Lee JE, Kim HS, Kwon YU, Jung GH, Lee CK, Yun HT, Kim CK. Responses of Photosynthetic Characters to Waterlogging in Soybean [Glycine max (L.) Merrill]. Korean J. Crop Sci 2010. 55: 111-118.
- 11. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001. 25: 402-408.
- 12. Scott HD, Deangelo J, Daniels MB, Wood LS. Flood duration effects on soybean growth and yield. Agron. J 1989. 81: 631-636.
- 13. Thomas AL, Guerreiro SMC, Sodek L. Aerenchyma formation and recovery from hypoxia of the flooded root system of nodulated soybean. Ann. Bot 2005. 96: 1191-1198.
- 14. Shi F, Yamamoto R, Shimamura S, Hiraga S, Nakayama N, Nakamura T, Yukawa K, Hachinohe M, Matsumoto H, Komatsu S. Cytosolic ascorbate peroxidase 2 (cAPX 2) is involved in the soybean response to flooding. Phytochem 2008. 69: 1295-1303.
- 15. Visser E, Cohen JD, Barendse G, Blom C, Voesenek L. An ethylene-mediated increase in sensitivity to auxin induces adventitious root formation in flooded Rumex palustris Sm. Plant Physiol 1996. 112: 1687-1692.
- 16. Lee JE, Kim HS, Kwon YU, Jung GH, Lee CK, Yun HT, Kim CK. Responses of root growth characters to waterlogging in soybean [Glycine max (L.) Merrill]. Korean J. Crop Sci 2010. 55: 1-7.