적요
This study was conducted to isolate a salt tolerant gene and to develop salt tolerant rice for reclaimed-saline areas through genetic transformation. A rice c/DRE binding factor 4 (OsCBF4) cDNA was isolated from rice (cv. Nipponbare) using RT-PCR. The full-length cDNA of the CBF4 gene consists of 1,429 nucleotides and 274 amino acid residues. The OsCBF4 shares from 33 to 49% identity of deduced amino acid sequence with other CBFs of rice. In order to develop salt tolerant rice, transgenic rice plants containing the OsCBF4 gene were obtained via Agrobacterium-mediated transformation. The stable incorporation of the OsCBF4 gene into rice genome was confirmed by PCR and Southern analysis. The stable expression of introduced gene was also validated by RT-PCR analysis in T2 plants. Biological assay of T3 progeny of the transgenic plants in Yoshida solution containing 120 mM Nacl for 2 weeks, confirmed that the OsCBF4 confers salt tolerance to transgenic rice plants. OsCBF4 transgene in the transgenic line CBF4-10 was markedly expressed up to over three-fold in the leaf by 120 mM NaCl treatment. Real-time PCR analysis revealed that the levels of the transgene expression were markedly increased under salt treatment. The transgenic line CBF4-10 which showed highest ability to recover from the saline stress could be used as a potential source for salt tolerance in rice breeding programs.
서 언
지구상의 염분지역 면적은 대륙표면의 약 6%에 해당하는 약 9억 5천만 ha로 지구상의 전체 작물재배 면적의 약 10% 에 해당한다(
Pessrakli 1994). 우리나라의 서남해안에 분포한 간척 가능 면적 중 농경지로 개발할 수 있는 면적은 약 402천 ha이며, 그 중 1999년 까지 117천 ha가 이미 간척되었으며, 현재 40천 ha의 새만금 간척지가 조성 중이어서, 염 토양은 계속적으로 증가하고 있다. 벼는 우리나라의 대표적 주곡작물 로 간척지에서 재배하기에 적합한 작물이나, 우리나라에서 재 배되는 자포니카형 벼는 내염성 유전자원이 보고되지 않았다. 다만 인디카형인 Pokkali 등이 내염성 벼로 보고되었으나, 자 포니카의 우량형질을 유지하면서 Pokkali의 염 저항성 관련 유전자만을 도입하여 자포니카형 내염성 벼를 개발하는 것은 교배육종법으로는 어려운 실정이다. 따라서 염분지역에서 재 배가 가능한 내염성 작물 개발은 간척지의 이용 효율 증진을 위해 효과적인 방법이라 할 수 있다.
생명공학기술을 이용한 내염성 작물 개발을 위해 항산화 관련 효소인 ascorbate peroxidase, glutathione reductase, superoxide dismutase, catalase 등을 과발현 시킨 형질전환 담배(
Allen et al. 1997;
Roxas et al. 1997), 유채(
Ashraf et al. 2008)에서 염 저항성이 보고되었다. C-repeat/dehydration-responsive element (c/DRE) binding factor (
CBF)는 애기장 대의 스트레스 유발 유전자 promoter부위에서 처음 분리되었 다(
Yamaguchi-shinozaki & Shinozaki 1994).
CBF 유전자 는 애기장대에서 과발현 시 고농도 염, 한발 및 저온에 대한 저항성을 증진하는 기능이 밝혀졌고(
Haake et al. 2002), 카 놀라(
Jaglo et al. 2001), 토마토(
Hsieh et al. 2002), 담배 (Kasuga et al. 2004) 등 작물에서도 재해저항성을 증진시켰 다. CBF1 형질전환 애기장대에서 내냉성이 향상되고(
Jaglo-Ottosen et al. 1998), 애기장대의
CBF2와
CBF3는 내한발성 과 내염성을 증진하였다(
Liu et al. 1998;
Nakashima et al. 2000). Dubouzet et al. (
2003)은 벼의
CBF3 유전자를 애기 장대에 형질전환하여 내염성, 내냉성, 내한발성이 증진된 식 물체를 개발하였고, Oh et al. (
2005)은 애기장대의
CBF3 유 전자를 벼에 도입하여 내염성, 내한발성, 내냉성 등 복합 내재 해성 형질전환 벼를 개발하였다. 벼에 보리의
CBF4 유전자 를 과발현 시킨 결과 한발, 고농도 염, 저온 스트레스에 대해 서 저항성이 증가되고(
Oh et al. 2007), 애기장대에서
CBF4 를 과발현 시킨 결과 저온과 한발 스트레스에 대한 저항성을 나타내었다(
Haake et al. 2002).
따라서 벼에서 분리한 CBF4 유전자의 염 저항성에 대한 역할을 확인하고 내염성 벼 유전자원 개발에 이용하고자 본 연구를 실시하였다.
재료 및 방법
유전자 분리
유전자 분리에는 Nipponbare 벼 품종을 이용하였고, CBF4 유전자를 분리하기 위하여 GenBank 데이터베이스에 등록된 애기장대와 벼의 CBF 유전자를 이용하여 해당부위의 염기서 열 구조를 분석한 후 각각의 프라이머쌍을 제작하여 RT-PCR 분석에 이용하였다. RT-PCR 반응은 One Step RNA PCR kit (Takara, Japan)을 이용하여 50°에서 역전사 반응을 30 분간 수행한 후, 94°에서 2분간 cDNA를 1회 변성시키고, 증폭반응(94° 30초, 60° 30초, 72° 1분 10초)을 30회 진 행한 후, 최종 신장 반응은 72°에서 7분간 수행 하였다. PCR 을 수행하여 증폭된 cDNA 단편을 Qiaquick Gel Extraction kit (Qiagen, USA)를 이용하여 정제하고, pGEM-T Easy vector (Promega, USA)에 클로닝한 후 JM109 E. coli 균주에 형질 전환하여 염기서열 분석에 이용하였다.
형질전환체 개발
OsCBF4 cDNA를 벼에 형질전환하기 위하여 ubiquitin promoter를 갖는 pSB22 벡터에 재조합하였고, 재조합된 pSB 2230 벡터는 freezing-thawing 방법을 이용하여
A. tumefaciens LBA4404에 도입하였다(
An 1987;
An et al. 1988). pSB2230 이 도입된
A. tumefaciens LBA4404를 AB 액체배지에서 진 탕 배양한 후, AAM 액체배지에 희석하고 동진벼의 캘러스를 접종하여 N
6 배지에서 3일간 공동배양하였다(
Lee et al. 1999). 공동배양한 캘러스는 5 mg/L phosphinothricin (PPT)이 첨 가된 1차 선발배지에서 3주간 배양한 후 6 mg/L PPT가 첨가 된 2차 선발배지에서 활발하게 증식하는 캘러스를 재분화배 지에 계대배양하여 재분화 식물체를 획득하였다(Table
1).
Table 1Media used for tissue culture and transformation of rice
Table 1
|
Medium |
Composition |
|
|
N6z callus induction |
N6 salts and vitamins, 30 g/L sucrose, 1 g/L casamino acid, 2 mg/L 2,4-D,2 g/L phytagel, pH 5.6 |
|
AB medium |
3 g/L K2HPO4, 1 g/L NaH2PO4, 1 g/L NH4Cl, 0.15 g/L KCl, 0.2 g/L MgSO47H2O, 0.012 g/L CaCl2•2H2O, 0.025 g/L FeSO4•7H2O, 5 g/L glucose, 4 mg/L tetracycline, pH 7.0 |
|
AAM medium |
AA salts and amino acids, MS vitamin, 68.5 g/L sucrose, 36 g/L glucose, 0.5 g/L casamino acid, 20 mg/L acetosyringone, pH 5.2 |
|
N6 co-culture |
N6 salts and vitamins, 30 g/L sucrose, 1 g/L casamino acid, 10 g/L glucose, 20 mg/L acetosyrin gone, 2 mg/L 2,4-D, 2 g/L phytagel, pH 5.2 |
|
First selection |
N6 salts and vitamins, 30 g/L sucrose, 1 g/L casamino acid, 5 mg/L PPT, 250 mg/L cefotaxime, 2 mg/L 2,4-D, 2 g/L phytagel, pH 5.6 |
|
Second selection |
N6 salts and vitamins, 20 g/L sucrose, 30 g/L sorbitol, 2 g/L casamino acid, 6 mg/L PPT, 250 mg/L cefotaxime, 2 mg/L 2,4-D, 0.5 mg/L BAP, 2 g/L phytagel, pH 5.6 |
|
Regeneration medium |
MSy salts and vitamin, 40 g/L sucrose, 20 g/L sorbitol, 125 mg/L cefotaxime, 0.1 mg/L NAA, 2.0 mg/L kinetin, 5 g/L phytagel, pH 5.6 |
|
Root induction |
MS salts and vitamins, 30 g/L sucrose, 125 mg/L cefotaxime, 2 g/L phytagel, pH 5.6 |
PCR 분석
형질전환 식물체의
OsCBF4 유전자 도입은 Wang 등(
1993) 의 방법에 따라 추출한 벼 genomic DNA와
CBF4 특이 primer쌍 [forward (GGATCCATGGAGGGGGG)와 reverse (CGAGCTCCTA ATAGGAGAAAAGGCTAA)]을 이용하 여 PCR을 수행하였다. Accupower PCR PreMix (Bioneer, Korea)를 이용하여 94°에서 2분간 변성반응을 거친 후, 증 폭반응(94° 30초, 65° 40초, 72° 50초)을 35회 진행하였 고, 최종 신장반응은 72°에서 7분간 수행하였다. PCR 산물 은 0.5 μg/ml EtBr이 포함된 1.0% agarose gel에서 50 V로 1시간 전기영동한 후 UV transilluminator에서 확인하였다.
Roger & Bendich (
1989)의 방법으로 genomic DNA 10 μg 을 이용하여
BamHI 제한효소를 16시간 0.8% agarose gel에 서 25 V, 16시간 전기영동한 후, Southern (
1975)의 방법을 이용하여 Hybond N
+ nylon membrane (Amersham, UK)에 DNA를 전이시켰다.
CBF4 cDNA를 Alkphos Direct Labelling kit (Amersham, UK)를 이용하여 탐침자를 제작하고, 잡종화 반응을 위해 membrane을 Alkphos Direct Detection kit를 이용하여 잡종화 과정을 수행한 후, X-ray film에서 3시간 노 출시켜 결과를 확인하였다.
Southern 분석을 통하여 OsCBF4 유전자 도입이 확인된 개체에서 도입 유전자의 발현을 확인하기 위하여 RT-PCR을 실시하였다. RT-PCR 반응은 잎에서 분리한 total RNA와 CBF4 특이 primer쌍 [forward (GTGTTTGTCAGGAGCG GAAGAA)와 reverse (TAA ACCCATCATCTCCCCCTTG)] 을 이용하여 PCR을 수행하였다.
내염성 생물검정 및 유전자 발현분석
동진벼와
OsCBF4 형질전환벼 T
3 세대 종자를 스포탁액으 로 소독하고 여지가 깔린 페트리디쉬에서 발아 시킨 후 유리 온실에서 Yoshida 양액(
Yoshida et al. 1976)을 이용하여 10 일간 어린 식물체를 육성하고 120 mM NaCl 조건에서 2주간 처리하였다. NaCl 처리 후, Yoshida 양액에서 1주일간 회복 시킨 후 농촌진흥청에서 설정한 내염성 달관기준표를 이용하 여 내염성 정도를 조사하였다.
Real-time PCR은 생물검정에서 선발된 계통의 도입유전 자 발현을 조사하기 위하여 실시하였다. T
3세대 종자를 10일 간 Yoshida 배양액(
Yoshida et al. 1976)으로 재배한 후, 60, 120 mM NaCl 조건에서 3일간 처리한 식물체의 잎과 뿌리에 서 tatal RNA를 분리하여 iScript™ Select cDNA Synthesis Kit (Bio-Rad, USA)로 cDNA를 합성하였다. Real-Time PCR 은 CFX96 (Bio-Rad, USA)를 이용하였고, iQ™SYBR Green Supermix와
CBF4 특이 primer쌍을 이용하여 95°에서 3분 을 거친 후 95°에서 20초, 63°에서 15초, 72°에서 20초 간 40회 반복하고 매 신장 단계에서 SYBR/FAM channel로 형광량을 측정하였다. 측정된 형광량을 이용한 유전자 발현량 분석은
Actin mRNA 발현량을 이용하여 각 처리구의
CBF4 mRNA 발현량에 대한 상대적인 양을 조사하였다.
결과 및 고찰
OsCBF4 cDNA는 1,429 bp 크기의 뉴클레오티드로 DNASIS 프로그램을 이용하여 염기서열을 분석한 결과 번역부위가 401 ~1,222 bp이며 AUG 개시코돈과 UAG 종지코돈이 존재하 고, 274개의 아미노산을 암호화하는 완전한 클론이었다(Fig.
1A).
Fig. 1.(A) The nucleotide and deduced amino acid sequences of OsCBF4. (B) Similarity of the deduced amino acid sequence of OsCBF4 to that of OsCBF1, OsCBF2, OsCBF3, OsCBF5, OsCBF6 and AtCBF1 synthase sequences, respectively.

OsCBF4 유전자는 벼의 다른
CBF 유전자와 33~49% 상 동성을 나타내었으나, 애기장대의
CBF1 유전자와는 상동성 이 낮았다. 벼의
CBF4는
CBF3와 49%의 가장 높은 상동성 을 나타내었으나 벼의 다른
CBF 유전자와는 30~44%의 낮 은 상동성을 보여 벼
CBF 유전자는 각각 서로 다른 경로를 통하여 유전자의 기능을 갖게 되었음을 시사하였다(Fig.
1B).
OsCBF4 벼 형질전환 식물체는 155개체를 획득하였고, OsCBF4 유전자 도입 여부를 확인하기 위하여, 특이 primer 쌍 을 이용하여 PCR을 실시한 결과 148개의 식물체에서 형질전환 에 이용된 pSB2230과 동일한 크기의 840 bp DNA 단편이 증 폭되어 OsCBF4 유전자의 벼 염색체내 삽입이 확인되었다.
OsCBF4 유전자 도입이 확인된 T
2세대 중 바스타액 검정 을 통하여 유전자 고정이 확인되고 GMO 포장에서 생육이 균일한 10계통을 무작위로 선발하여
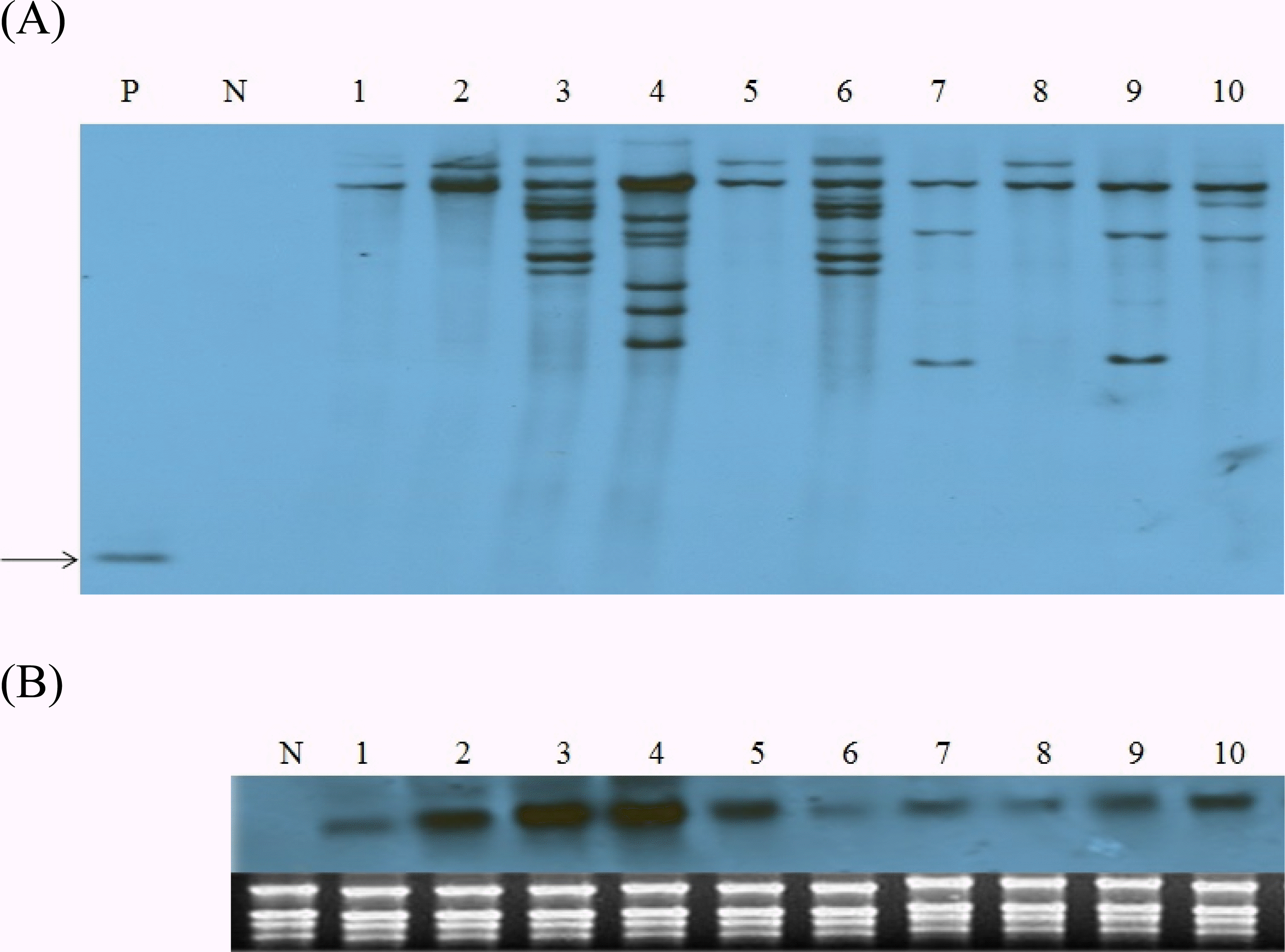
CBF4 유전자 도입 조성 을 Southern blot 분석을 통하여 확인하였다. DNA 조성 분 석 결과 비 형질전환 벼(lane N)에서는 어떠한 단편도 검출되 지 않았으나, 분석에 이용된 모든 형질전환체에서 pSB2230 유래
OsCBF4 유전자가 1~7사본이 벼 게놈에 도입되었음을 확인하였다(Fig.
2A).
Fig. 2.Molecular characterization of transgenic rice lines expressing OsCBF4. A. Southern blot analysis. Genomic DNAs in lanes P and 1 to 10 were digested with enzymes BamHI which is unique in the T-DNA region. Arrow indicates the fragment (840 bp) hybridized with the OsCBF4 cDNA probe. B. Northern blot analysis of the expression of OsCBF4 transgene. Total RNAs from leaf samples of the same lines in (A) were analyzed. P, pSB2230 plasmid vector; N, nontransgenic Dongjin; lanes 1-10, independent transgenic lines.
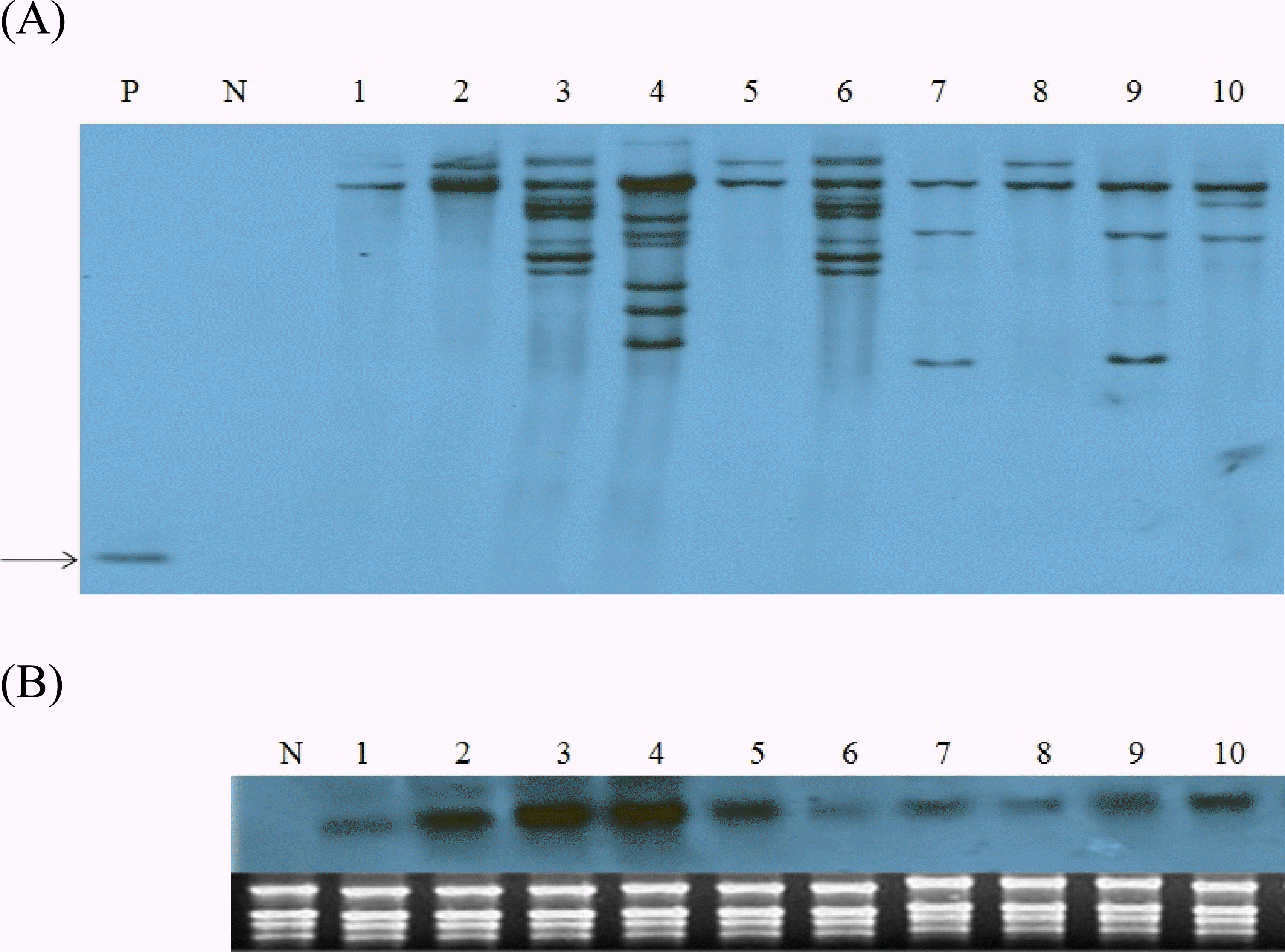
Southern 분석에서
OsCBF4 유전자 도입이 확인된 동일 계통에 대한 전이 유전자의 발현은 northern blot 분석으로 확인하였다. 모든 형질전환 계통에서
OsCBF4 유전자가 강하 게 발현되었으며 특히 CBF4-2, CBF4-3, CBF4-4는 RNA 밴 드의 발현이 강하였으나 불임이 심하였고, 비 형질전환체에서 는
OsCBF4 유전자의 발현이 확인되지 않았다(Fig.
2B). 이 것은
OsCBF4 유전자가 형질전환 벼의 게놈에 안정적으로 도입되었을 뿐만 아니라, 도입된 유전자는 벼 고유의
CBF 유 전자와 상호작용에 의한 억제 작용 없이 상시 발현을 조절하는 ubiquitin promoter에 의해 정상적으로 전사체가 합성되어진 결과이다.
OsCBF4 유전자의 정상적 발현이 확인된 형질전환 계통 중 종자의 불임이 없고 농업적 형질이 양호한 계통의 T
3 종자를 이용하여 염 스트레스에 대한
OsCBF4 유전자의 발현을 유 묘 생물검정을 통하여 확인하였다. 10일간 자란 어린 식물체 를 120 mM NaCl 양액에서 2주간 염 스트레스를 준 후 1주 일간 염이 제거된 양액에서 회복시킨 결과, 비 형질전환 벼(동 진)는 대부분 고사하였다(Fig.
3A). 하지만
OsCBF4 유전자 도입 형질전환 벼는 많은 개체에서 염 스트레스에 의한 생리적 장애로 부터 회복되어 정상적으로 생육하였고, 특히 CBF4-10 계통은 72.5%의 높은 회복을 나타내었다(Fig.
3A,
B). 이상 의 결과는 보리의
CBF4 유전자 과발현 형질전환 벼가 건조, 저온, 고 농도의 염 스트레스 조건에서 저항성이 증가하였다 는 연구와 유사한 결과이다(
Oh et al. 2007).
Fig. 3.Biological assays of the NaCl-treated OsCBF4 transgenic rice plants. Non-transgenic and transgenic plants were grown in Yoshida solution containing 120 mM NaCl for 2 weeks and transferred to the same solution containing no NaCl for 1 week for recovery. A. Symptoms of the non-transgenic (middle) and transgenic (left and right) plants. B. Survival of Germination of non-transgenic (NT) and transgenic (CBF4-6 and CBF4-10) plants under salt stress. aTotal number of plants used in each assay. bNumber of survival plants. cPercent of survived plants (survival/total ×100).

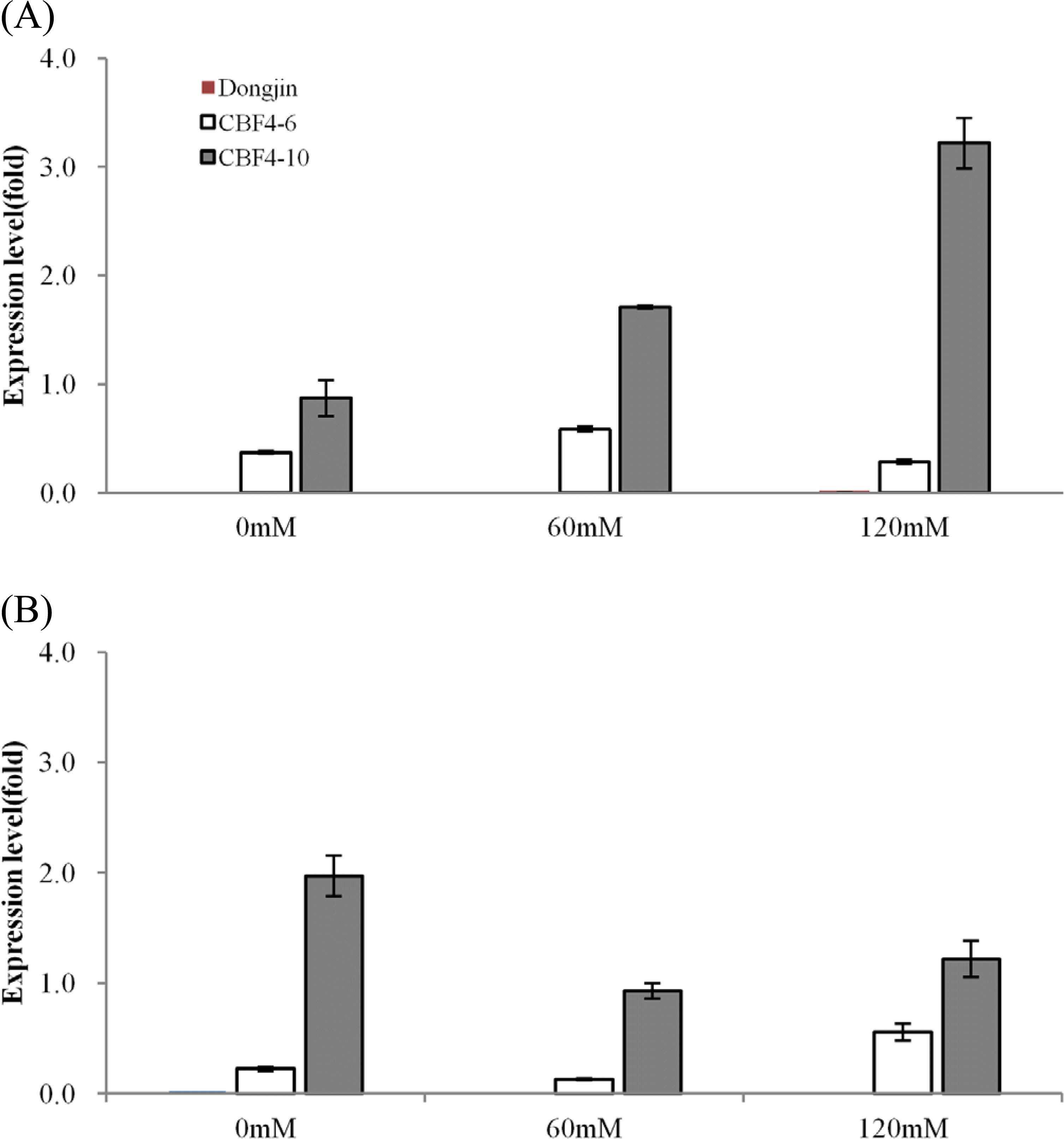
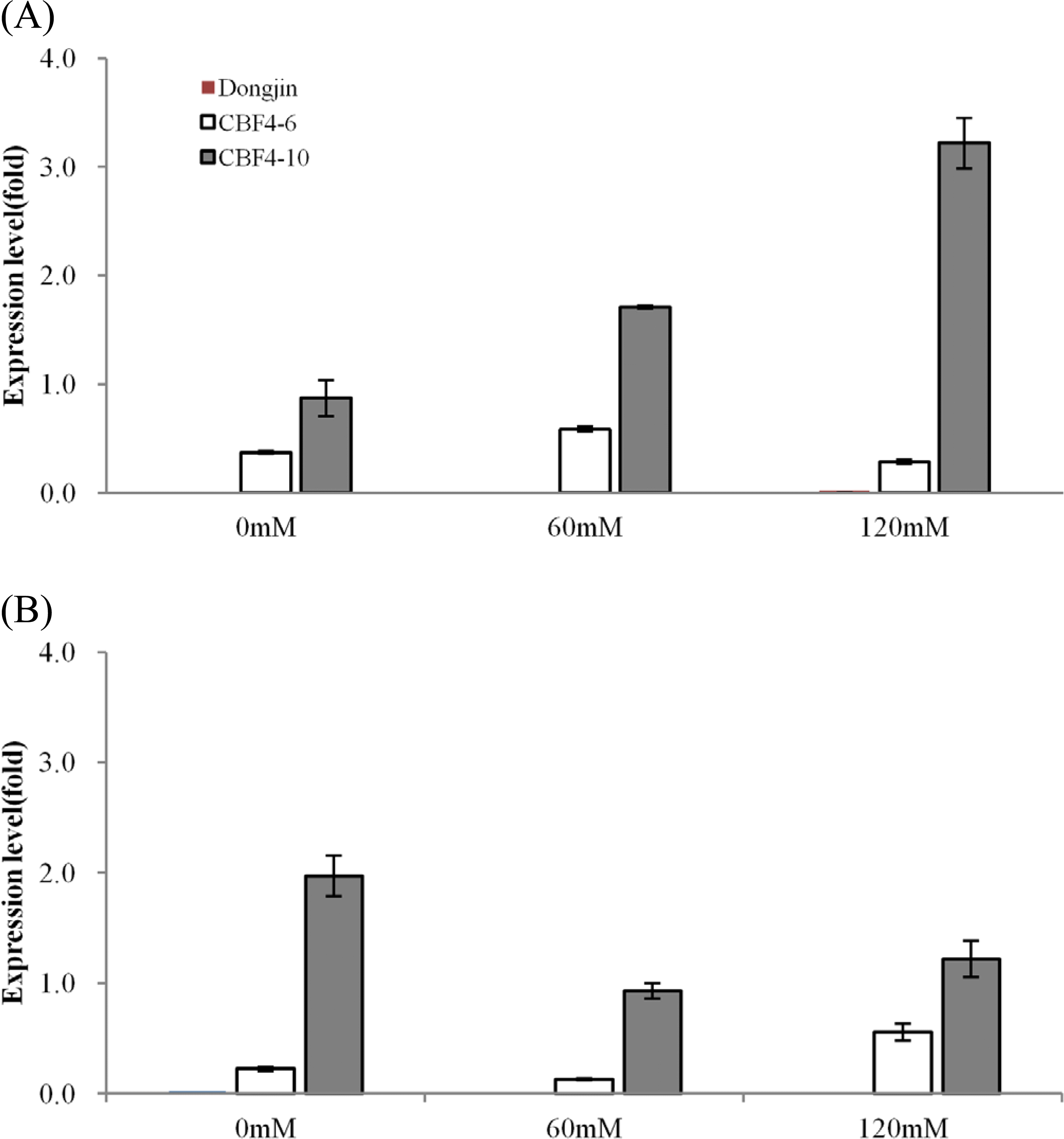
내염성 생물검정에 이용한 동일 형질전환 계통의 어린 식 물체에 농도별 NaCl을 3일간 처리한 후 전이 유전자의 발현 을 real-time PCR로 측정한 결과,
CBF4 전사체가 비 형질전 환 벼의 잎과 뿌리에서는 전혀 검출되지 않았고, 형질전환 계 통에서는 모든 처리에서 검출되었다. CBF4-6 계통은 NaCl 처리와 무처리 모든 구의 잎과 뿌리에서 전사체 발현량이 큰 차이를 나타내지 않았으나, 염 스트레스 처리 후에도 많은 개 체에서 정상적인 생육을 나타낸 CBF4-10 계통은 잎에서는 염 처리 농도가 증가할수록 전사체 발현량이 약 3배 이상 증 가하였고 뿌리에서는 반대로 발현량이 1/2 수준으로 감소하 였다(Fig.
4A,
B). 이것은 염 스트레스 조건에서 지상부에서 의 염의 독성 회피와 삼투 포텐셜의 불균형을 극복하기 위한 전사조절 유전자인
CBF 유전자의 적극적 발현의 결과임을 시 사한다(
Sakuma et al. 2006). 내염성 벼 개발에 이용된
OsCBF4 유전자는 스트레스 유도성 promoter가 아닌 상시 발현하는 ubiquitin에 의해 조절되어 전이 유전자는 항시적으로 발현되 었다. 하지만 염 스트레스에 의해
OsCBF4 유전자의 발현이 3배 이상 증가하는 결과를 보였다. 이것은 상시 발현 35S promoter에 의해 조절되는 형질전환 식물체에서 고온, 저온, 상 처 등 여러 종류의 스트레스에 의해 도입된 GUS유전자의 발현 이 증가하였다는 Maghuly et al. (
2008)과 ubiquitin promoter에 의해 조절되는 PR10 유전자 도입 형질전환 벼에서 고 농도의 염 스트레스에서 처리 시간별 도입 유전자의 발현 정도가 변 화하였다는 Kim et al. (
2010)의 결과와 유사하였다.
Fig. 4.Expression of the OsCBF4 transgene in the leaf (A) and root (B) of NaCl-treated non-transgenic and transgenic rice plants. Relative mRNA levels in plants of CBF4-6 (white bars) or CBF4-10 (gray bars) or control line (Dongin) (red bars) were determined by quantitative PCR. Transcription of the OsCBF4 transgene was examined under 0, 60 and 120 mM NaCl treatment for 3 days.
OsCBF4 유전자 과발현 형질전환 계통은 종자 발아시 비 형질전환 벼에 비해 다소 지연되는 결과를 보였는데 이것은 Dubouzet et al. (
2003)의 형질전환 종자의 발아와 유묘 생장 이 지연되었다는 보고와 유사하다. Liu et al. (
1998)은
CBF3, ABF3 유전자가 과발현된 형질전환 애기장대에서 심각한 생 육 장애를 나타낸다고 하였다. 하지만,
OsCBF4 형질전환 벼 는 GMO 포장재배에서 일부 계통에서 불임에 의해 수량이 감소되기도 하였으나 많은
CBF4 형질전환 계통은 일반 벼와 유사한 생육과 정상적인 임성을 나타내었다. 이상의 결과는
CBF 유전자 과발현 형질전환 벼에서 왜소화 증상 없이 정상 적으로 생육하였다는 Oh et al. (
2005,
2007)과 유사한 결과 이다.
이상의 결과를 종합하면, CBF4 형질전환 벼는 고 농도의 염 스트레스 조건에서 무처리 보다 전이 유전자의 발현량이크게 증가할수록 내염성 정도는 높아졌다. 즉, 전이 유전자의 발현량과 식물체의 염 저항성간에 관계가 있음을 의미한다. 따라서, OsCBF4 유전자는 염에 대한 저항성 기능을 가지고 있으며, OsCBF4 형질전환 벼 계통은 내염성 유전자원으로 활용할 가치가 있는 것으로 판단된다.
적 요
내염성과 관련 있는 벼의 CBF4 유전자(OsCBF4)를 벼에서 과발현 시켜 염 스트레스 조건에서 저항성이 증진된 내염성 유전자원을 개발하였다. RT-PCR을 수행하여 분리한 OsCBF4 유전자는 1,429 bp 크기의 뉴클레오티드로 274개의 아미노 산으로 구성되었고, 벼의 다른 CBF 유전자와 33~49% 상동 성을 나타내었다. 형질전환 식물체는 PCR과 Southern분석으 로 OsCBF4 유전자의 벼 게놈 내 도입을 확인하였다. 전이 유전자는 벼 게놈 내에 1~4사본이 도입되었고, 선발된 형질 전환 계통 모두에서 전이 유전자가 강하게 발현되었다. 염 스트 레스 조건에서 형질전환 식물체는 비 형질전환 벼 보다 생육 정도가 양호하였으며, 특히 CBF4-10 계통은 염 처리 후 많은 식물체가 살아남았다. Real-time PCR 분석 결과 CBF4-10 계 통은 120 mM NaCl 처리 시 전이 유전자 OsCBF4 전사체 발 현이 잎에서 무처리 대비 약 3배 이상 증가하였다. 결론적으 로 일반 벼에 도입된 OsCBF4 전이 유전자는 내염성 증진 기 능이 있으며, OsCBF4 전이 유전자의 발현이 높은 형질전환 벼 계통은 내염성 벼 육종 소재로 이용할 수 있을 것으로 평가 된다.
사 사
본 성과물은 농촌진흥청 연구사업(레스베라트롤 생산벼 익 산526의 농업형질 비교분석 및 특성평가. PJ009528032014) 의 지원에 의해 이루어 진 것임.
References
- 1. Allen RD, Webb RP, Schake SA. Use of transgenic plants to study antioxidant defenses. Free Radic. Biol. Med 1997. 23: 473-479.
- 2. An G. Binary Ti vectors for plant transformation and promoter analysis. Methods Enzymol 1987. 153: 292-305.
- 3. An G, Ebert PR, Mitra A, Ha SB, Schilperoort R, Verma DP. Binary vectors. Plant Mol. Manual 1988. A3: 1-19.
- 4. Ashraf M, Ali Q. Relative memberane permeability and activities of some antioxidant enzymes as the key determinants of salt tolerance in canola (Brassica napus L.). Environ. Exp. Bot 2008. 63: 266-273.
- 5. Chu CC, Wang CC, Sun CS, Hsu C, Yin KC, Chu CY, Bi FY. Establishment of an efficient medium for anther culture of rice through comparative experiments on the nitrogen sources. Sci. Sin 1975. 18: 659-668.
- 6. Dubouzet JG, Sakuma Y, Ito Y, Kasuga M, Dubouzet EG, Miura S, Seki M, Shinizaki K, Yamaguchi-Shinozaki K. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in droght-,high-salt- and cold-responsive gene expression. Plant J 2003. 33: 751-763.
- 7. Haake V, Cook D, Riechmann JL, Pineda O, Thomashow MF, Zhang JZ. Transcription factor CBF4 is a regulator of drought adaptation in Arabidopsis. Plant Physiol 2002. 130: 639-648.
- 8. Hsieh TH, Lee JT, Charng YY, Chan MT. Tomato plants ectopically expressing Arabidopsis CBF1 show enhanced resistance to water deficit stress. Plant Physiol 2002. 130: 618-626.
- 9. Jaglo-Ottosen KR, Gilmour SJ, Zarka DG, Schabenberger O, Thomashow MF. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science 1998. 280: 104-106.
- 10. Jaglo KR, Kleff S, Amundsen KL, Zhang X, Haake V, Zhang JZ, Deits T, Thomashow MF. Components of the Arabidopsis C-repeat/dehydration-responsive element binding factor cold-response pathway are conserved in Brassica napus and other plant species. Plant Physiol 2001. 127: 910-917.
- 11. Kasuga M, Miura S, Shinozak K, Yamaguchi-Shinozaki K. A combination of the Arabidopsis DREB1A gene and stress-inducible rd29A promoter improved drought and low temperature stress tolerance in tobacco by gene transfer. Plant Cell Physiol 2004. 45: 346-350.
- 12. Kim HJ, Baek SH, Shin WC, Seo CS, Park MR, Ko JK, Yun SJ. Development of salt-tolerant transgenic rice using soybean PR10 gene. Korean J. Breed. Sci 2010. 42: 540-546.
- 13. Lee S, Jeon JS, Jung KH, An G. Binary vectors for efficient transformation of rice. J. Plant Biol 1999. 4: 310-316.
- 14. Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought and low temperature responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998. 10: 1391-1406.
- 15. Maghuly F, Khan MA, Fernadez EB, Druart P, Watillon B, Laimer M. Stress regulated expression of the Gus-marker gene (uidA) under the control of plant calmodulin and viral 35S promoters in a model fruit tree rootstock: Prunus incisa×serrula. J. Biotech 2008. 135: 105-116.
- 16. Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Plant Physiol 1962. 15: 473-497.
- 17. Nakashima K, Shinwan ZK, Sakuma Y, Seki M, Miura S. Organization and expression of two Arabidopsis DREB2 genes encoding DRE-binding proteins involved in dehydration-and high-salinity-responsive gene expression. Plant Biotechnol 2000. 42: 657-665.
- 18. Oh SJ, Song SI, Kim YS, Jang HJ, Kim SY, Kim M, Kim YK, Nahm BH, Kim JK. Arabidopsis CBF3/ DREB1A and CBF3 in transgenic rice increased tolerance to abiotic stress without stunting growth. Plant Physiol 2005. 138: 341-351.
- 19. Oh SJ, Kwon CW, Choi DW, Song SI, Kim K. Expression of barley HvCBF4 enhances tolerance to abiotic stress in transgenic rice. Plant Biotechnol 2007. 5: 646-656.
- 20. Pessarakli M. Handbook of plant and crop stress 1994. 697. New York, Marcel Dekker, Inc.
- 21. Roger SO, Bendich AJ. Gelvin SB, Schilperoort RA, Verma DPS. Extraction of DNA from plant tissues. Plant Molecular Biology Manual 1989. A6: 1-10.
- 22. Roxas VP, Smith RK, Allen ER, Allen RD. Overex- pression of glutathione S-transferase/glutathione peroxidase enhances the growth of transgenic tobacco seedlings during stress. Nat. Biotechnol 1997. 15: 988-991.
- 23. Sakuma Y, Maruyama K, Osakabe Y, Qin F, Seki M, Shinozaki K. Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell 2006. 18: 1292-1309.
- 24. Southern EM. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol 1975. 98: 503-517.
- 25. Wang H, Qi M, Cutler AJ. A simple method of preparing plant samples for PCR. Nucleic Acids Res 1993. 21: 4153-4154.
- 26. Yamaguchi-Shinozaki K, Shinozaki K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, high-salt stress. Plant Cell 1994. 6: 251-264.
- 27. Yoshida S, Forno DA, Cock JH, Gomez KA. Laboratory manual for physiological studies of rice 1976. 3rd edition. Philippines, The international rice research institute. 61-66.