적요
Low-molecular-weight glutenin subunits (LMW-GS) play a crucial role in the processing quality of wheat flour. They are encoded multi gene family located at the Glu-A3, Glu-B3 and Glu-D3 on the short arm of chromosome 1A, 1B and 1D respectively. Typical LMW-GSs are composed of three parts including a short N-terminal domain, a relatively short repetitive domain and a C-terminal domain. Further, typical LMW-GS sequences are divided into LMW-s, LMW-m and LMW-i types, on the basis of the first amino acid of the mature proteins (serine, methionine and isoleucine, respectively). Although it is known that the allelic variation of LMW-GSs affect the properties of dough, it is still not clear which LMW-GSs confer better bread-making quality because of the larger number of expressed subunits and their overlapping mobility with abundant gliadin proteins. Therefore, it is important to characterize LMW-GS genes and develop functional markers to identify different LMW-GS alleles for application in wheat breeding. In this review, we discuss the various aspects of LMW-GS, including their structural characteristics, the development of marker, relationship between LMW-GSs and bread wheat quality, and genetic engineering of the LMW-GSs.
서 언
밀은 대략 BC 7500-6500년 전부터 경작된 인류 최초의 작 물이다. BC 6000-5000년 사이에 육배체 밀(
T. aestivum) 과 엠머 밀(emmer wheat,
T. timopheevi var. araraticum와
T. turgidum var. dicocoides)이 메소포타미아와 서이란 지역에 서 관개농업을 통해 경작되었다는 기록이 있다(
Feldman 1995). 밀은 귀리나 보리와 같이 밀족(
Triticeae)에 속하고 벼과 (
Poaceae family)에서 가장 크고 중요한 속 중의 하나이다(
Dewey 1984). 밀족은 일반적으로
Aegilops,
Elymus,
Hordeum,
Secale,
Triticum 5개의 속으로 구성된다(
Löve 1984,
Stebbins 1956). 밀속과 밀족에서 일반적으로 염색체수는 x = 7이다. 야생 이배체 밀
Triticum monococcum (AA 염색체),
T. tauschii (DD 염색체) 와
T. speltoides (SS) 에서는 2n = 2x = 14개, 사배체 밀
T. turgidum (AABB 염색체)와
T. timopheevii (AAGG 염색체)는 2n = 4x = 28개이다(
Feldman 1976). 보통밀
T. aestivum (AABBDD 염색체)은 6배체 밀로서, 2n = 6x = 42개의 염색 체를 가진다(Zohary & Hopf 1993).
밀 저장 단백질이 빵 만들기에 가장 중요한 역할을 한다고 알려져 있으며(
Morrison 1988,
Wall 1979), 빵 가공성은 정 성적으로 특정 단백질의 유무 및 특정 단백질 서브유닛의 유 무가 밀접한 관련성이 있다고 알려져 있다(
Gupta et al. 1989,
Johansson et al. 1998 and 2001,
Payne et al. 1987). 밀가루 가 빵, 국수, 과자와 같은 다양한 가공적성을 가지는 것은 전 적으로 글루텐 단백질들에 기인한다(
Weegels et al. 1996). 260여 년 전 Beccari에 의해 밀 글루텐 단백질 추출이 보고된 이후, 밀 글루텐 단백질의 추출방법에 따른 글루텐 단백질의 분류 와 글루텐 구조가 반죽에 미치는 기능적 특성을 밝히는 연구가 꾸준히 이어져오고 있다(
Payne 1987,
Shewry et al. 2002,
Wieser 2007). 글루텐 단백질은 알콜 수용액 [60-70% (v/v) ethanol또는 50% (v/v) propan-1-ol]의 용해도에 따라 녹는 글리아딘과 녹지 않은 글루테닌으로 나눠진다(
Wieser 2007). 글리아딘은 시스테인 잔기가 없어 이황화 결합이 없거 나, 있어도 분자 내 이황화 결합(intra-disulfide bond)를 형성 하며 단량체 형태로 존재한다.
밀 저장 단백질은 글루테닌과 글리아딘의 두개의 주된 단 백질로 구성되어 있다. 글루테닌은 고분자 글루테닌 서브유닛 [high-molecular-weight glutenin subunit (HMW-GS)]과 저 분자글루테닌 서브유닛 [low-molecular-weight glutenin subunit (LMW-GS)]으로 구성되며, HMW-GS는 1A, 1B, 1D 염색체 장완에 있는
Glu-A1,
Glu-B1,
Glu-D1에 의해 암호화되고 (
Payne et al. 1980), LMW-GS는 1A, 1B, 1D 염색체 단완 에 있는
Glu-A3,
Glu-B3 그리고
Glu-D3에 의해 암호화된다 (
Gupta & Shepherd 1990,
Jackson et al. 1996). 밀 품종에서 HMW-GS와 LMW-GS의 유전적 변이가 밀 반죽의 물성의 변화 및 가공성에 영향을 미친다고 보고되고 있다(
D’Ovidio & Masci 2004,
Nagamine et al. 2000).
국민 1인당 밀 소비량이 연간 32.4 kg으로 쌀 다음으로 높 지만, 최근 이상기후와 바이오 연료용 곡물 수요의 증가로 인한 국제 곡물 가격 상승으로 자급률이 2% 이하인 국내 현실에서 밀 자급률 향상은 어느 때 보다 절실한 실정이다(농업통계연 감 2011). 국내산 밀의 자급률 향상을 위해 숙기 단축과 수량성 및 재해 저항성 증진에 대한 연구가 활발히 진행되는 한편으 로, 밀의 가공적성에 많은 영향을 미치는 글루테닌의 기능 연구 를 위하여 국내 밀품종에 대한 SDS-PAGE나 STS (sequence tagged sites) 분자표지를 이용한 판별이 이루어지고 있다(
Park et al. 2011).
본 총설에서는 글루텐 복합체(Gluten complex) 중에서도 가공적성에 많은 영향을 미치고 있지만 분석상의 어려움 때 문에 연구가 미진한 저분자글루테닌 서브유닛의(LMW-GS)의 분자 구조, 기능(quality property), 육종 현장에서 활용도가 높은 분자표지 마커 및 형질전환에 관해 기술하고자 한다.
저분자 글루테닌 서브유닛 및 기본 구조
LMW-GS를 조절하는 유전자는 1번 염색체 단완(short-arm) 에 위치해 있으며, 이들 유전자좌는 각각
Glu-A3,
Glu-B3및
Glu-D3으로 명명되었다(Singh & Shepherd 1988). 이러한
Glu-3유전자는 γ및 ω글리아딘 발현에 관여하는
Gli-1 유전 자좌와 관련이 되어 있는 것으로 알려져 있다(
Jackson et al. 1996). SDS-PAGE분석을 통한 LMW-GS 발현은,
Glu-A3 유전자좌에서 6개의 패턴으로 알려져있고,
Glu-B3 유전자좌 에서는 9개의 패턴이,
Glu-D3 유전자좌에서 5개의 패턴으로 발현되어 전체적으로 20개의 LMW-GS 단백질이 발현되는 것으로 알려져 있다(
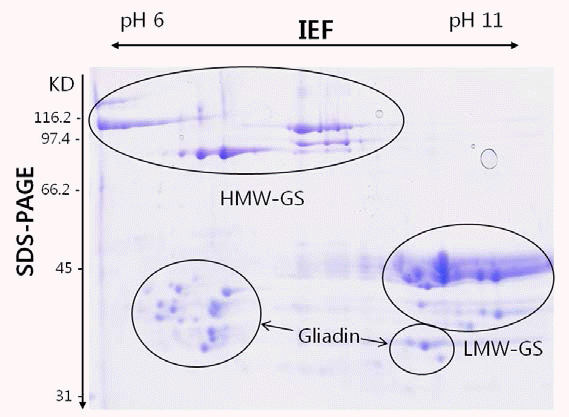
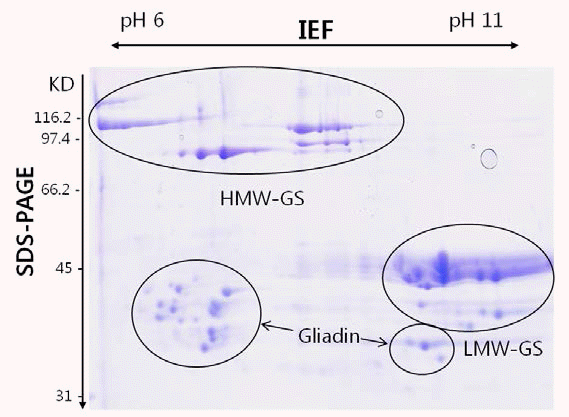
Gupta & Shepherd 1990). 글루테닌 단 백질은 이차원전기영동 즉, 분자량과 등전점(isoelectric point, pI)에 따라 A, B, C 세 그룹으로 나뉘는데 A 그룹은 HMWGS, B 그룹은 전형적인 LMW-GS, C 그룹은 LMW-GS와 α/ β, γ 글리아딘의 혼합 그룹으로 존재한다(Fig.
1). 분자간 이 황화결합을 형성하는 시스테인 잔기의 위치 및 N-말단과 C- 말단의 아미노산의 배열에 따라 12 그룹으로 분류하기도 한 다(
Ikeda et al. 2002,
Juhász & Gianibelli 2003). 또한, LMW-GS는 성숙 단백질(mature protein)의 N-말단의 첫 아 미노산 잔기에 따라 LMW-i (이소루신), LMW-m (메치오닌) 그리고 LMW-s (세린)의 세 종류로 나뉜다(
Cloutier et al. 2001,
Lew et al. 1992). 이들 세 종류의 LMW-GS 단백질은 이차원 전기영동과 N-말단아미노산 서열분석에 의해 확인되었 다(
Ikeda et al 2006,
Lew et al. 1992,
Maruyama-Funatsuki et al. 2004,
Masci et al. 1998).
Fig. 1.Two-dimensional gel electrophoresis (IEF × SDS-PAGE) of glutenin fraction of wheat cultivar Chinese Spring. The HMW-GS, LMW-GS and gliadin fractions are indicated.
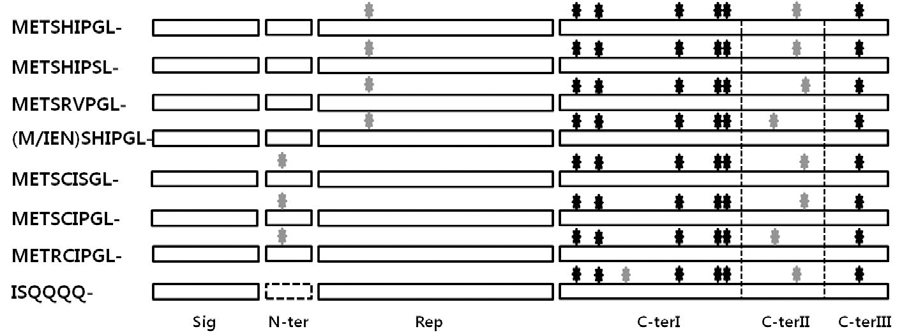
전형적인 LMW-GS의 구조는 Fig.
2와
3, Table
2에 나타 낸 바와 같이 20개 아미노산의 signal peptide, N-말단 부분, 비교적 짧은 서열의 반복 도메인 그리고 C-말단으로 구성되 어 있으며 i-타입 LMW-GS의 경우 N-말단 부분이 다른 타입 의 LMW-GS들과 달리 바로 ISQQQQ-의 서열로 시작한다. LMW-s 타입은 모든 밀 품종들에서 가장 발현량이 많으며 평균 분자량(35,000-45,000 daltons)도 LMW-m타입(30,000- 40,000 daltons)보다 크다(
Lew et al. 1992,
Masci et al. 1998,
Tao & Kasarda 1989). LMW-s 타입의 N-말단 아미노산 서 열은 현재까지 SHIPGL- 하나만 보고된 반면, LMW-m 타입은 METSHIPGL-, MENSHIPGL-, METSHIPSL-, METSRVPGL-, METSRIPGL-, 그리고 METSCIPGL-로 다양하다(
Ikeda et al. 2002 and
2006,
Kasarda 1989,
Lew et al. 1992,
Masci et al. 1998,
Tao & Kasarda 1989). Table
2에
Glu-A3,
Glu-B3 및
Glu-D3 유래의 LMW-GS 단백질의N-말단 서열에 관하여 정리하였다.
Ikeda et al. (2006)은 LMW-s타입의 SHIPGL- 서열은 LMW-m 타입의 METSHIPGPL- 서열이 asparaginyl peptidase에 의해 전사후변형(post-translational modification) 으로 인해N-말단의 3개 잔기가 절단되었다고 제시하고 있다. 이와 유사한 현상이 오메가 글리아딘에서도 보고되었다(
DuPont et al. 2004).
Fig. 2.Diagrams representing the structures of typical LMW-GSs as deduced amino acids from their genes. The grey and black stars indicate cysteins involved in inter-molecular and intra-molecular bonds, respectively. Sig; signal peptide, N-ter; N-terminal domain; Rep; repetitive domain and C-ter; C-terminal domain.
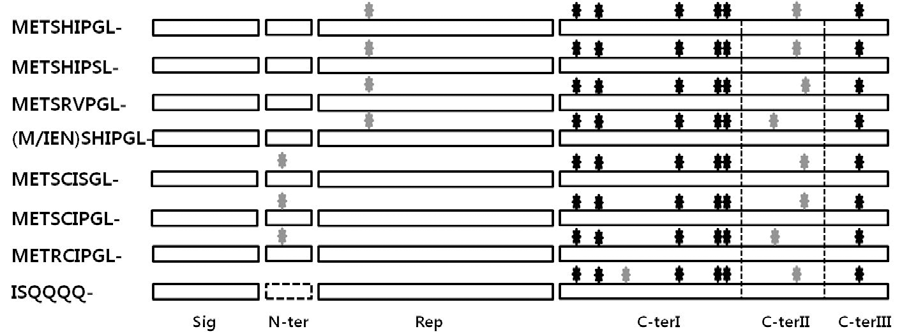
Fig. 3.Alignments of deduced amino acid sequences of Glu-A3, Glu-B3 and Glu-D3allelic variants. Identical residues are shaded. The position of cystein residues are marked with filled down arrow. Sig; signal peptide, N-ter; N terminal domain; Rep; repetitive domain and C-ter; C-terminal domain and these positions are marked with filled nabla. GluD3-1 specific hydrophobic residues are underlined. GluA3-1 specific glutamine repeats are also underlined.

Table 2.N-terminal amino acid sequences of the deduced Glu-A3, Glu-B3 and Glu-D3 proteins.
Table 2.
|
Locus |
Genes |
N-terminal amino acid sequence Type based on the first amino acid |
Type (Group) based on Ikeda et al. |
Reference |
|
|
Glu-A3
|
GluA3-1
|
ISQQQQ(Q/P/A)P- |
LMW-i |
VI (11 &12) |
Wang et al. 2010
|
|
GluA3-2
|
GluA3-2 |
LMW-m |
IV(6) |
|
GluA3-3z
|
ISQQQQQ- |
LMW-i |
|
|
|
Glu-B3
|
GluB3-1
|
MENSHIPGL- |
LMW-m |
II(3) |
Wang et al. 2009
|
|
GluB3-2
|
MENSHIPGL- |
LMW-m |
II(3) |
|
GluB3-3
|
MENSHIPGL- |
LMW-m |
II(3) |
|
GluB3-4
|
METSHIPSL- |
LMW-m |
I (2) |
|
|
Glu-D3
|
GluD3-1
|
METSRVPGL- |
LMW-m |
III (5) |
Zhao et al. 2007
|
|
GluD3-2
|
METRCIPGL- |
LMW-m |
V (10) |
|
GluD3-3
|
(M/IEN)SHIPGL- |
LMW-s |
II (4) |
|
GluD3-4
|
METSCISGL- |
LMW-m |
IV (7) |
|
GluD3-5
|
METSHIPGL- |
LMW-m |
I (1) |
|
GluD3-6
|
METSCIPGL- |
LMW-m |
IV (8 & 9) |
LMW-GS는 분자 내 그리고 분자 간 이황화결합을 형성하 는 8개의 시스테인을 포함하며 분자 간 이황화결합을 형성하 는 시스테인의 위치에 따라 2개로 분류된다(Fig.
2, Fig.
3). LMW-m, LMW-s 타입은 N-말단 부분 또는 반복 도메인 위 치에 시스테인 잔기가 1개, C-말단에 나머지 1개가 존재하며 LMW-i 타입은 C-말단에 분자 간 이황화결합을 형성하는 2 개의 시스테인 잔기가 존재한다. 또한 Fig.
3의 밑줄 부분에 나타낸 바와 같이 i-타입 LMW-GS (
GluA3-1)의 경우 C- 말 단 I 부분에 6개의 글루타민의 반복이 보이며, METSRVPGL- 로 시작하는
GluA3-1는 반복도메인에 PVIILQQ와 같은 소 수성 잔기들이 존재하였다. 이와 같이 반복 도메인에 반복 모 티프의 삽입 또는 결실 그리고 분자간 이황화 결합을 형성하 는 시스테인 잔기의 위치 차이가 기능의 차이와 관련이 되어 있을 것으로 추론하고 있다(
Ikeda et al. 2002,
Wieser 2007).
LMW-GS 유전자 및 유전자 좌 마커 연구
현재 일반적으로
Glu-A3,
Glu-B3 그리고
Glu-D3 유전자 좌에 대한 STS 마커가 개발되어 있지만, 육종 현장에서는
Glu-A3 (
Wang et al. 2010),
Glu-B3 (
Wang et al. 2009) 마 커만 사용하고 있다(http://maswheat.ucdavis.edu/protocols/ FunctionalMarkers/FM_quality.htm#wang2009b). Table
1에 나타낸 것처럼
Glu-D3 유전자 좌 마커는 특정
Glu-D3 유전자 만을 구분하기 때문에 아직 범용으로는 사용되지 못하고 있다. 유전자 좌특이 마커는 지금까지
Glu-A3 위치에는 7개(
a,
b,
c,
d,
e,
f,
g),
Glu-B3 위치에는 9개(
a,
b,
c,
d,
e f,
g,
h,
i) 그리고
Glu-D3 위치에는 5개(
a,
b,
c,
d,
e,
f ) 등 21개의 유 전자 좌(SDS-PAGE mobility에 의한 분류)에 대해 확인하여 육종 현장에 많이 사용하고 있다(
Gupta & Shepherd 1990,
Jackson et al. 1996,
Zhang et al 2004). 또한 새로운 LMWGS 유전자 좌를 발견하고 마커화 하려는 시도가 보고되고 있 다(
McIntosh et al. 2008).
Wang et al. (2010)은 밀 품종 Aroona (
GluA3c)와 6개의 Aroona 근동질 계통(
gluA3a,
b,
d,
e,
f)과 밀 품종Glenlea (
GluA3g)를 이용하여
Glu-A3 위치 에서 유전자 특이 프라이머로
GluA3-1,
GluA3-2, 그리고
GluA3-3의 전장 open reading frame (ORF)를 나타내는 유 전자들을 분리하였고, SDS-PAGE 분류법에 의한
gluA3a,
gluA3b,
gluA3ac (
Glu-A3a와
c를 구별),
gluA3d,
gluA3e,
gluA3f,
gluA3g 유전자 좌 구별이 가능한 프라이머를 제작하 였으며, 또한
Glu-A3b +
Glu-A3f,
Glu-A3d +
Glu-A3f,
Glu-A3d +
Glu-A3f 그리고
Glu-A3b +
Glu-A3e를 구별할 수 있는 다중 검출 마커(multiplex marker)를 개발하여 141 개 CIMMYT 밀 품종에서 검증하였다.
Glu-B3 위치에서는 8 개의 Aroona (
Glu-B3b)와Aroona 근동질 계통(
Glu-B3a,
c,
d,
f,
g,
h,
i)과 밀 품종Cheyenne (
Glu-B3e)를 이용하여 유전 자 특이 프라이머로
GluB3-1,
GluB3-2,
GluB3-3 그리고
GluB3-4 완전한 ORF를 지닌 유전자들을 분리하였고, 위의 9개 유전자 좌 구분을 위한
gluB3a,
gluB3b,
gluB3c,
gluB3d,
gluB3e,
gluB3fg (
Glu-B3f와 g를 구별),
gluB3g,
gluB3h,
gluB3i,
gluB3bef (
Glu-B3b,
e,
f를 구별)유전자 좌 특이 프라 이머를 제작하였고 161개의 품종 및 후대라인에서 검증하였 다(
Wang et al. 2009).
Glu-D3 위치에서는 Chinese Spring (
Glu-D3a), BT2288A (
Glu-D3e), Silverstar (
Glu-D3b), Sunco (
Glu-D3b), Aroona (
Glu-D3c), Norin 61 (
Glu-D3d), Tasman (
Glu-D3a), Hartog (
Glu-D3e)의 8개의 보통 밀 품 종을 이용하여 유전자 특이 프라이머로
GluD3-11,
GluD3-12,
GluD3-21,
GluD3-22,
GluD3-23,
GluD3-31,
GluD3-32,
GluD3-41,
GluD3-42,
GluD3-43,
GluD3-5,
GluD3-6의 12 종류의 유전자를 분리하였고, 이중
GluD3-5와
GluD3-6는 모든 품종에서 이들 유전자를 공통으로 가지고 있는 것을 확 인하였다. 또한
GluD3-21/22,
GluD3-22,
GluD3-23,
GluD3-31,
GluD3-32, GluD3-41, GluD3-43에 특이적인 STS를 개발하 여
Glu-D3a,
b,
c 그리고 f를 가지는 중국 밀 20개 품종에서 검증하였다(
Zhao et al. 2006 and
2007). Table
1에서는
Glu-3 각각의 유전자 좌를 구별하기 위한 STS 마커 프라이 머를 표시하였고, Fig.
3에서
Glu-A3,
Glu-B3 그리고
Glu-D3 유전자들의 유추 아미노산을 나타내었다.
Table 1.Allele-specific PCR markers for the discrimination of Glu-3 alleles defined by protein mobility in common wheat.
Table 1.
|
Locus |
Marker/ Target gene |
Sequence (5’-3’) |
Allele |
Expected fragment size (bp) |
Chromosome |
Reference |
|
|
Glu-A3
|
gluA3a
|
LA1F: AAACAGAATTATTAAAGCCGG SA1R: GGTTGTTGTTGTTGCAGCA |
Glu-A3a
|
529 |
1AS |
Wang et al. 2010
|
|
gluA3b
|
LA3F: TTCAGATGCAGCCAAACAA SA2R:GCTGTGCTTGGATGATACTCTA |
Glu-A3b
|
894 |
|
|
|
gluA3d
|
LA3F: TTCAGATGCAGCCAAACAA SA4R: TGGGGTTGGGAGACACATA |
Glu-A3d
|
967 |
|
|
|
gluA3e
|
LA1F: AAACAGAATTATTAAAGCCGG SA5R: GGCACAGACGAGGAAGGTT |
Glu-A3e
|
158 |
|
|
|
gluA3f
|
LA1F: AAACAGAATTATTAAAGCCGG SA6R: GCTGCTGCTGCTGTGTAAA |
Glu-A3f
|
552 |
|
|
|
|
gluA3g
|
LA1F: AAACAGAATTATTAAAGCCGG SA7R:AAACAACGGTGATCCAACTAA |
Glu-A3g
|
1345 |
|
|
|
|
gluA3ac
|
LA1F: AAACAGAATTATTAAAGCCGG SA3R: GTGGCTGTTGTGAAAACGA |
Glu-A3a Glu-A3c
|
573 |
|
|
|
|
|
Glu-B3
|
gluB3a
|
SB1F: CACAAGCATCAAAACCAAGA SB1R: TGGCACACTAGTGGTGGTC |
Glu-B3a
|
1095 |
1BS |
Wang et al. 2009
|
|
gluB3b
|
SB2F: ATCAGGTGTAAAAGTGATAG SB2R: TGCTACATCGACATATCCA |
Glu-B3b
|
1570 |
|
|
|
gluB3c
|
SB3F: CAAATGTTGCAGCAGAGA SB3R: CATATCCATCGACTAAACAAA |
Glu-B3c
|
472 |
|
|
|
gluB3d
|
SB4F: CACCATGAAGACCTTCCTCA SB4R: GTTGTTGCAGTAGAACTGGA |
Glu-B3d
|
662 |
|
|
|
gluB3e
|
SB5F: GACCTTCCTCATCTTCGCA SB5R: GCAAGACTTTGTGGCATT |
Glu-B3e
|
669 |
|
|
|
gluB3g
|
SB7F: CCAAGAAATACTAGTTAACACTAGTC SB7R: GTTGGGGTTGGGAAACA |
Glu-B3g
|
853 |
|
|
|
gluB3h
|
SB8F: CCACCACAACAAACATTAA SB8R: GTGGTGGTTCTATACAACGA |
Glu-B3h
|
1022 |
|
|
|
gluB3i
|
SB9F: TATAGCTAGTGCAACCTACCAT SB9R: TGGTTGTTGCGGTATAATTT |
Glu-B3i
|
621 |
|
|
|
gluB3bef
|
SB10F: CATCAACAACAAATAGTACTAGAA SB10R: GGCGGGTCACACATGACA |
Glu-B3b Glu-B3e
|
750 |
|
|
|
gluB3fg
|
SB6F: TATAGCTAGTGCAACCTACCAT SB6R: CAACTACTCTGCCACAACG |
Glu-B3f Glu-B3g
|
812 |
|
|
|
|
Glu-D3
|
GluD3-21/22
|
M2F12: TTGGGCCTAATCGCTCGC M2R12: TAGTCTCCATCTGCGCAATT |
Glu-D3b Glu-D3a
|
884 |
1DS |
Zhao et al. 2007
|
|
GluD3-22
|
M2F2: CTCGTCTTTGCCCTCCTCA M2R2: CTAAACAACGGTGACCCAAT |
Glu-D3b? Glu-D3a?
|
958 |
|
|
|
GluD3-23
|
M2F3: TCTGTACTTTGTGTGTGATCG M2R3: ACTGCTGCTGGAGGAATAG |
Glu-D3f Glu-D3c
|
725 |
|
|
|
GluD3-31
|
M3F1:ACAAGTGCCATTGCACAAATG M3R1: GATAGATGGATGAACAAATA |
Glu-D3a Glu-D3b
|
528 |
|
|
|
GluD3-32
|
M3F2: CAAGTGCCATTGCACAAATT M3R2: AATGATGGTTGTTGCGGTAT |
Glu-D3f Glu-D3c
|
334 |
|
|
|
GluD3-41
|
M4F1: AAGTAGTTAGCACCAATCCAT M4R1: CCTGTTGTTGTTGTTGTTGTT |
Ndz
|
773 |
|
|
|
GluD3-43
|
M4F3:GCATCAAAACCAAGCAAAAG M4R3: GGCTGAACAATAGGGATTTA |
Ndz
|
413 |
|
|
LMW-GS 기능 연구
LMW-GS 단백질이 HMW-GS 보다 많이 발현되고 함량 도 많지만, 가공적성에 미치는 영향에 대해서는 연구가 미흡 하고, 발현되는 각 단백질에 대한 연구도 밝혀진 것이 거의 없는 실정이다. 그 원인으로 SDS-PAGE에 의해 분리되는 LMW-GS 양상이 글리아딘과 유사한 양상을 보여 분자량에 따른 동정이 어렵기 때문이다(
Appelbee et al. 2009,
D’Ovidio & Masci 2004). 이차원 전기영동(2DE) 분석 방법은 1차원 SDS-PAGE 보다 훨씬 많은 정보를 제공하므로 LMW-GS 발현 단백질의 정성 및 정량 분석으로 보다 정밀하고 다양한 자료를 제시할 수 있으며, 최근에는 2차원 전기영동과 MALDITOF- MS (matrix-assisted laser desorption / ionization time-of-flight mass spectrometry)와 같은 펩타이드 질량분 석법의 결합을 통한 LMW-GS 분석이 실시되고 있다(
Liu et al. 2010). 또한, 최근 글루테닌 관련 유전자서열 특이적 분자 표지를 활용한 분석이 많이 이루어지고 있으며 (
Gale 2005), 육종의 효율성을 높이고자
Glu-1및
Glu-3의 다중검출 PCR 마커를 활용하는 연구가 수행되고 있다 (
Liang et al. 2010,
Salmaowicz & Moczulski 2004,
Sui et al. 2010,
Wang et al. 2010, Zhang et al. 2008).
HMW-GS와 제빵적성과의 관계는 폭넓게 연구되었다. 예 를 들어 HMW-GS 5 + 10 단백질이 HMW-GS 2 + 12에 비 해 좋은 제빵적성을 가진다고 보고되었다(
Payne et al. 1980 and
1987). 하지만 LMW-GS의 제빵적성에 관련한 연구는 아직 시작단계에 불과하다. 최근의 연구들에서 LMW-GS도 HMW-GS 만큼 제빵적성에 중요한 역할을 한다는 결과들이 보고되고 있는데, 초강력분인 Canadian Western Extra-Strong (CWES)의 특정 LMW-GS를 일본 밀 품종에 도입한 근동질 계통에서 밀반죽의 물성이 강화되었다(
Takata et al. 2001). 또한 CWES와 일본 밀 품종의 재조합 계통들에서 KS2로 명 명된 42kDa의 LMW-GS가 비슷한 크기의 LMW-GS인 HS1 에 비해 제빵 적성에서 더 좋은 결과를 보였고, HMW-GS 5 + 10과 KS2, HMW-GS 5 + 10과 HS1 그리고 HMW-GS 2 + 12과 KS2, HMW-GS 2 + 12과 KS2의 조합 실험에서도 HMW-GS 5 + 10과 KS2라인에서 가장 좋은 결과를 보였는 데, 이는 특정 LMW-GS 단백질이 밀반죽 물성에 많은 영향을 미 친다는 것을 보여주고 있다(
Maruyama-Funatuski et al. 2004).
일반적으로 LMW 글루테닌 구성성분들이(allelic composition) 이 보통밀에서 반죽강도(dough strength) 및 신장성 (extensibility)에 영향을 미친다고 알려져 있으며, 이것은 LMW-GS와 HMW-GS가 중합체를 형성하고, 이 중합체가 단량체인 글리아딘과의 결합하여 글루텐 복합체(Gluten complex) 를 형성하고 밀반죽 물성(dough rheological property)에 영 향을 미친다고 보고되었다(
Gupta et al. 1989 and
1994,
Nieto-Taladriz et al. 1994,
Payne et al. 1987,
Ruiz & Carrillo 1995,
Sontag-Strohm et al. 1996).
Glu-3 위치에서 특정 LMW-GS 유전자 좌가 반죽강도 그리고 신장성에 영향을 미 치는 것이 밝혀졌는데, 여러 문헌에서
Glu-A3,
Glu-B3 및
Glu-D3 유전자 좌와 밀반죽 품질(dough quality effects)의 관계를 정립하였다(
Gupta et al. 1989,
1990,
1991 and
1994,
Metakovsky et al. 1990). Rmax (maximum dough resistance) 값은
Glu-A3 유전자 좌에서 b>d>e>c,
Glu-B3 유전자 좌에 서 i>b=a>e=f=g=h>c 그리고
Glu-D3에서 e>b>a>c>d의 순 서를 보였다. 신장성 부분에서는
Glu-B3 bbb (각각
Glu-A3,
Glu-B3 및
Glu-D3)의 조합일 때 최고의 값을 보이고, bbc 또 한 좋은 결과 값을 보였다(
Cornish et al. 1993).
중국과 호주 공동연구 그룹에서 특정 LMW-GS 유전자 좌 (allele)의 기능 연구를 위해 호주 밀 품종인 Aroona에 5개의
Glu-A3 (b, c, d, e, f) 라인, 9개의
Glu-B3 (a, b, c, d, e, f, g, h, i)라인 그리고 5개의
Glu-D3 (a, b, c, d, f) 근동질 계통 을 이용하여 밀반죽의 품질인자(quality parameter)를 측정하 였는데 이 중
Glu-A3d,
Glu-B3b,
Glu-B3g,
Glu-B3i 라인들 에서 좋은 제빵 품질을 보였고,
Glu-D3 유전자 좌들에서는 미미한 결과를 보였다(
Zhang et al. 2012). 이러한 결과들을 볼 때 국내에서 우수한 제빵 적성을 가진 밀 품종 육성을 위 해서는 상기의 LMW-GS 유전자 좌의 도입이 시급할 것으로 생각된다.
LMW-GS단백질의 구조, 유전자 좌 분석, 유전자 발현과 밀가루 반죽의 물성과의 관계에서 탄성에 관여하는 메카니즘 등을 연구하는 주된 이유는 밀의 가공적성을 개량하는데 있 다. 지금까지의 연구는 특정 밀 품종에서 글루테닌과 글리아 딘의 정성적인 분석과 기능과의 관계를 연구하는데 집중되어 있었다면 앞으로는 분석 기술의 발달과 함께 밀 반죽 물성에 많은 영향을 미치는 정량적인 요인 즉, 글루테닌 단백질의 절 대량(glutenin quantity), 글리아딘 단백질의 절대량(gliadin quantity), 글리아딘 단백질과 글루테닌 단백질의 구성비 (gliadin / glutenin ratio), 고분자 글루테닌 단백질과 저분자 글루테닌 단백질의 구성비(HMW-GS / LMW-GS ratio)와 같은 요소들과 기능과의 관계를 분석하는 연구에 집중하여야 할 것이다(
Veraverbeke et al. 2002).
밀 LMW-GS의 형질전환 연구
LMW-GS가 밀 글루텐 복합체 에서 많은 역할을 하고 밀 가루 가공적성의 중요한 결정인자 임에도 이들의 분자구조와 기능에 관한 연구는 아직 미미하다. 이러한 이유로 밀의 가공 적성 개량 및 밀가루 반죽에서의 기능 연구를 위해 안정적이 고 재현성 있는 밀 형질전환 기술을(
Vasil et al. 1992,
Weeks et al. 1993)을 이용하여 여분의 LMW-GS 유전자를 추가적 으로 도입하는 시도가 진행되어 왔다.
Masci et al. (2003)은 밀 품종 Cheyenne에서
Glu-D3 유래의 5’ 및3’ 인접서열 (flanking sequence)을 포함하는 1,200 bp와 1,600 bp 두 유 전자를 분리하여 보통 밀인 Bob White에 유전자 총(Biolistic transformation)을 이용하여 과발현 형질전환체를 육성하였 고, Southern blot과 2DE로 유전자 삽입과 도입 LMW-GS 단백질 발현을 확인하였다. 기능분석을 위해 형질전환체 밀가 루와 비형질전환체의 밀가루에서 SDS 침강법 실험을 한 결 과 비형질전환체의 밀가루가 형질전환체 보다 높은 값을 보 였는데 이것은 글루텐 단백질의 양 보다 특정 LMW-GS가 기능에 영향을 미친다는 것을 나타내고 있다.
Araki et al. (2008)은 HMW-GS 1By9와 i 타입 LMW-GS (이케다 분류 법에 의한 그룹 11)을 벼 글루테린 돌연변이체인 LGC1에 아 그로박테리움법으로 형질 전환하였고 1차원 및 2차원 전기영 동으로 HMW-GS와 LMW-GS 단백질 발현을 확인하였다. 또한 각각의 형질전환체를 교배하여 HMW-GS와 LMW-GS 모두 벼 저장 단백질 중합체에 분자간 이황화결합에 의해 삽 입되었음을 보여주었다.
현재 일반적으로 LMW-GS 유전자는
Glu-A3 위치에서
GluA3-1,
GluA3-2,
GluA3-3의 3종류
Glu-B3 위치에서는
GluB3-1,
GluB3-2,
GluB3-3,
GluB3-4 등 4종류 그리고
Glu-B3 위치에서는
GluD3-1,
GluD3-2,
GluD3-3,
GluD3-4,
GluD3-5,
GluD3-6의 6종류의 유전자가 존재하며, 각각의 위 치(loci)에서 아미노산의 삽입, 결실, 변이에 의해 차이를 보 이게 되며, 각
Glu-A3,
Glu-B3 및
Glu-D3 유전자 좌는 2~3 개의 유전자와 Pseudo gene으로 구성된다(
Wang et al. 2009,
Wang et al. 2010,
Zhao et al. 2006 and
2007). 이들 각 유 전자들의 구조에 따른 기능적 차이를 보기 위해서는 이들 각 종류의 유전자별로 과발현 형질전환체를 제작하여 밀 반죽 물성을 확인해보거나, 대장균이나 효모, 곤충을 이용하여 발 현시킨 재조합 단백질(
Gali 1989,
Tamas & Shewry 2006)을 base flour에 혼합하여 물성의 변화를 분석하는 등의 협력연 구를 통해 분자적 수준에서의 LMW-GS 단백질의 기작과 기 능 분석 연구가 이루어져야 한다. 이러한 연구를 통해 축적된 자료를 기반으로 하여 강력분 및 가공적성 맞춤형 밀 품종 육 종이나 형질전환체 육성이 이루어져야 소비자 맞춤형 밀 생 산 및 다양성이 있는 식량 자원 확보에 기여를 할 것이다.
결 론
밀가루에서 저분자 글루테닌 서브유닛(LMW-GS)의 유전 적 변이가 고분자 글루테닌 서브유닛(HMW-GS) 만큼 밀 반 죽의 물성의 변화 및 가공적성에 많은 영향을 미친다. 총 글 루테닌 단백질의60 % 정도를 차지하는 LMW-GS는 밀반죽 의 신장성에 주로 관여하며, HMW-GS와 분자간 이황화 결 합을 통해 중합체를 형성하여 탄성에 영향을 미치며, 또한 단 량체인 글리아딘과 글루텐 중합체를 형성하여 밀반죽 특유의 물성을 나타낸다. 하지만, LMW-GS가 많은 기능을 가졌음에 도 불구하고 다수의 유전자(multi-gene)으로 존재하고 글리아 딘과의 혼재성으로 인해 유전자 좌 분석과 개별 단백질의 기 능 연구에 어려움이 있지만, 많은 수의 LMW-GS유전자에 대 한 연구가 진행되어 유전자서열 특이적 분자표지를 활용한 분석이 많이 이루어지고 있다. LMW-GS의 기능 연구를 위해 근동질 계통이나 재조합 계통을 이용하여 특정 유전자 좌와 빵 가공성(bread-making quality)에 관한 분석이 이루어졌으 며, 특정 LMW-GS 유전자의 기능 분석을 위해 형질전환체 분석이나 재조합발현을 통한 분석에 의한 연구가 진행되고 있다. 또한 정성적인 기능 분석과 더불어 글리아딘 단백질과 글루테닌 단백질의 비율 또는 HMW-GS와 LMW-GS의 비율 과 같은 정량적인 분석 결과와 기능과의 상관관계를 해명하 는 연구도 진행되어야 할 것이다.
사 사
본 연구는 농촌진흥청 국립농업과학원 농업과학기술 연구 개발사업(과제번호: PJ010116)의 지원에 의해 이루어진 것임.
References
- Andersen JR, Lubberstedt T. Functional markers in plants. Trends Plant Sci 2003. 8: 554-560.
- Appelbee MJ, Mekuria GT, Nagasandra V, Bonneau JP, Eagles HA, Eastwood RF. Novel allelic variants encoded at the Glu-D3 locus in bread wheat. J. Cereal Sci 2009. 49: 254-261.
- Araki E, Ikeda TM, Ohgihara Y, Toyoda A, Yano H. Development of transgenic rice (Oryza sativa L.) expressing wheat high- and low-molecular-weight glutenin subunit proteins. Korean J. Breed. Sci 2008. 58: 121-128.
- Bietz JA, Wall JS. Isolation and characterization of gliadin-like subunits from glutenin. Cereal Chem 1973. 50: 537-547.
- Branlard G, Dardevet R, Saccomano F, Lagoutte F, Gourdon J. Genetic diversity of wheat storage proteins and bread wheat quality. Euphytica 2001. 119: 59-67.
- Branlard G, Dardevet M, Amiour N, Igrejas G. Allelic diversity of HMW and LMW glutenin subunits and omega-gliadins in French bread wheat. Genet. Resour. Crop Evol 2003. 50: 669-679.
- Cloutier S, Rampitsch C, Penner GA, Lukow OM. Cloning and expression of a LMW-i glutenin gene. J. Cereal Sci 2001. 33: 143-154.
- Cornish GB, Békés F, Allen HM, Martin JM. Flour proteins linked to quality traits in an Australian doubled haploid wheat population. Aust. J. Agric. Res 2001. 52: 1339-1348.
- Cornish GB, Burridge PM, Palmer GA, Wrigley CW. Mapping the origins of some HMW and LMW glutenin subunit alleles in Australian germplasm. Proceedings of the 42nd Australian Cereal Chemistry Conference. Sydney 1993. 00: 255-260.
- DeweyDR. The genomic system of classification as a guide to intergeneric hybridisation with the perennial Triticeae. Gene manipulation in plant improvement. 1984. New York: Plenum Publ. Crop.
- D'Ovidio R, Masci S. The low-molecular-weight glutenin subunits of wheat gluten. J. Cereal Sci 2004. 39: 321-339.
- DuPont FM, Vensel W, Encarnacao T, Chan R, Kasarda DD. Similarities of omega gliadins from Triticum urartu to those encoded on chromosome 1A of hexaploid wheat and evidence for their post-translational processing. Theor. Appl. Genet 2004. 108: 1299-1308.
- FeldmanM. Wheats. In: Simmonds NW Evoliution of crop plants. England: Longman Group Limited, (Ed). 1976. 00: pp. 120-128.
- FeldmanM. Wheats. In: Smartt J, Simmonds NW. Evolution of crop plants. Harlow, UK: Longman Scientific and Technical, (Eds). 1995. pp. 185-192.
- Flaete NES, Uhlen AK. Association between allelic variation at the combined Gli-1, Glu-3 loci and protein quality in common wheat. J. Cereal Sci 2003. 37: 129-137.
- Gale KR. Diagnostic DNA markers for quality traits in wheat. J. Cereal Sci 2005. 41: 181-192.
- Galili G. Heterologous expression of a wheat high molecular weight glutenin gene in Escherichia coli. Proc. Natl. Acad. Sci. USA 1989. 86: 7756-7760.
- Gianibelli MC, Larroque OR, MacRitchie F, Wrigley CW. Biochemical, genetic, and molecular characterization of wheat glutenin and its component subunits. Cereal Chem 2001. 78: 635-646.
- Gupta RB, Shepherd KW. Two-step one-dimensional SDS-PAGE analysis of LMW subunits of glutenin. I. Variation and genetic control of the subunits in hexaploid wheats. Theor. Appl. Genet 1990. 80: 65-74.
- Gupta RB, Singh NK, Sheperd KW. The cumulative effects of allelic variation in LMW and HMW glutenin subunits on physical dough properties in the progeny of two bread wheats. Theor. Appl. Genet 1989. 77: 57-64.
- Gupta RB, Paul JG, Cornish GB, Palmer GA, Bekes F, Rathjen AJ. Allelic variation at glutenin subunit and gliadin loci, Glu-1, Glu-3 and Gli-1, of common wheats. 1. Its additive and interaction effects on dough properties. J. Cereal Sci 1994. 19: 9-17.
- GuptaRB. BekesF. WrigleyCW. Predicting values of LMW glutenin alleles for dough quality of bread wheat. In: Bushuk W, Tkachuk R. Gluten proteins. St. Paul. Minnesota. USA: American Association of Cereal Chemists, (Eds). 1990. pp. 615-621.
- Gupta RB, Bekes F, Wrigley CW. Prediction of physical dough properties from glutenin subunit composition in bread wheats: correlation studies. Cereal Chem 1991. 68: 328-333.
- Ikeda TM, Araki E, Fujita Y, Yano H. Theor. Appl. Genet 2006. 112: 327-334.
- Ikeda TM, Nagamine T, Fukuoka H, Yano H. Theor. Appl. Genet 2002. 104: 680-687.
- Jackson EA, Morel MH, Sontag-Strohm T, Branlard G, Metakovsky Radaelli R. Proposal for combining the classification systems of alleles of Gli-1 and Glu-3 loci in bread wheat. J. Genet Breed 1996. 50: 321-336.
- Juhász A, Gianibelli MC. Information hidden in the low molecular weight glutenin gene sequences. The Gluten Proteins 2003. In: Proc 8th Gluten Workshop; Bitervo, Italy. pp 62-65.
- Johansson E, Svensson G. Variation in bread-making quality: effects of weather parameters on protein concentration and quality in some Swedish wheat cultivars grown during the period 1975-1996. J. Cereal Sci. Food Agr 1998. 78: 109-118.
- KasardaDD. Glutenin structure in relation to wheat quality. In: Pomeranz Y Wheat is unique. St. Paul. Minnesota. USA: American Association of Cereal Chemists, (Ed). 1989. pp. 277-302.
- Lew EJL, Kuzmicky DD, Kasarda DD. Characterization of low molecular weight glutenin subunits by reversedphase high-performance liquid chromatography, sodium dodecylsulfate-polyacrylamide gel electrophoresis, and Nterminal amino acid sequencing. Cereal Chem 1992. 69: 508-515.
- Liang D, Tang J, Peña RJ, Singh R, He X, Shen X, Yao D, Xia X, He Z. Characterization of CIMMYT bread wheats for high and low molecular weight glutenin subunits and other quality-related genes with SDS-PAGE, RP-HPLC and molecular markers. Euphytica 2010. 172: 235-250.
- Liu L, Ikeda TM, Branlard G, Peña RJ, Rogers WJ, Lerner SE, Kolman MA, Xia X, Wang L, Ma W, Appels R, Yoshida H, Wang A, Yan Y, He Z. Comparison of low molecular weight glutenin subunits identified by SDS-PAGE, 2-DE, MALDI-TOF-MS and PCR in common
wheat. BMC Plant Biol 2010. 10: 124-148.
- Löve A. Conspectus of the Triticeae. Feddes Repertorium 1984. 95: 425-521.
- Maruyama-Funatsuki W, Takata K, Nishio Z, Tabiki T, Yahata E, Kato A, Saito K, Funatsuki H, Saruyama H, Yamauchi H. Identification of lowmolecular-weight glutenin subunits of wheat associated with bread-making quality. Plant Breed 2004. 123: 355-360.
- Masci S, D'Ovidio R, Lafiandra D, Kasarda DD. Characterization of a low-molecular-weight glutenin subunit gene from bread wheat and the corresponding protein that represents a major subunit of the glutenin polymer. Plant Physiol 1998. 118: 1147-1158.
- Masci S, D'ovidio R, Scossa F, Patacchini C, Lafiandra D, Anderson OD, Blechl AE. Production and characterization of a transgenic bread wheat line overexpressing a low-molecular-weight glutenin subunit gene. Mol. Breed 2003. 12: 209-222.
- McIntoshRA. YamazakiY. DubcovskyJ. RogersJ. MorrisC. SomersDJ. AppelR. DevoKM. Catalogue of gene symbols for wheat. Proceedings of the 11th international wheat genetics symposium 2008. 24-29 August 2008; Brisbane, Qld, Australia.
- Metakovsky EV, Wrigley CW, Bekes F, Gupta RB. Gluten polypeptides as useful genetic markers of dough quality in Australian wheats. Aust J. Agric Res 1990. 41: 289-306.
- MorrisonWR. Lipids. In: Pomeranz Y Wheat: chemistry and Technology. St. Paul. USA: AACC, (Ed). 1988. pp. 373-439.
- Nagamine T, Kai Y, Takayama T, Yanagisawa T, Taya S. Allelic variation at the Glu-1 and Glu-3 loci in southern Japanese wheats, and its effects on gluten properties. J. Cereal Sci 2000. 32: 129-135.
- Nieto-Taladriz MT, Perretant MR, Rousset M. Effect of gliadins and HMW and LMW subunits of glutenin on dough properties in the F6 recombinant inbred lines from a bread wheat cross. Theor. Appl. Genet 1994. 88: 81-88.
- Park CS, Kang CS, Jeung JU, Woo SH. Influence of allelic variations in glutenin on the quality of pan bread and white salted noodles made from Korean wheat cultivars. Euphytica 2011. 180: 235-250.
- Payne PI, Law CN, Mudd EE. Control by homologous group 1 chromosomes of the high-molecularweight subunits of glutenin, a major protein of wheat
endosperm. Theor. Appl. Genet 1980. 58: 113-120.
- Payne PI, Seekings JA, Worland AJ, Jarvis MG, Holt LM. Allelic variation of glutenin subunits and gliadins and its effect on bread making quality in wheat: analysis of F5 progeny from Chinese Spring. J. Cereal Sci 1987. 6: 103-118.
- Payne PI. Genetics of wheat storage proteins and the effect of allelic variation on breadmaking quality. Annu. Rev. Plant Physiol 1987. 38: 141-153.
- Ruiz M, Carrillo JM. Separate effects on gluten strength of Gli-1 and Glu-3 prolamine genes on chromosomes 1A and 1B in durum wheat. J. Cereal Sci 1995. 21: 137-144.
- Salmanowicz BP, Moczulski M. Multiplex polymerase chain reaction analysis of Glu-1 high molecular glutenin genes from wheat by capillary electrophoresis with laser-induced flourescence detection. J. Chromato 2004. 1032: 313-318.
- Shan X, Clayshulte SR, Haley SD, Byrne PF. Variation for glutenin and waxy alleles in the US hard winter wheat germplam. J. Cereal Sci 2007. 45: 199-208.
- Shewry PR, Halford NG, Tatham AS. High molecular weight subunits of wheat glutenin. J. Cereal Sci 1992. 15: 105-120.
- Shewry PR, Halford NG, Belton PS, Tatham AS. The structure and properties of gluten: an elastic protein from wheat grain. Philosophical Transactions of the Royal Society of London. Series B. Biological Sciences 2002. 357: 133-142.
- Shewry PR, Halford NG, Tatham AS, Popineau Y, Lafiandra D, Belton PS. The high molecular weight subunits of wheat glutenin and their role in determining wheat processing properties. Adv. Food Nutr. Res 2003. 45: 219-302.
- Singh NK, Shepherd KW, Cornish GB. A simplified SDS-PAGE procedure for separating LMW subunits of glutenin. J. Cereal Sci 1991. 14: 203-208.
- Sontag-Strohm T, Payne PI, Saovaara H. Effect of allelic variation of glutenin subunits and gliadins on baking quality in the progeny of two biotypes of bread wheat cv. Ulla. J. Cereal Sci 1996. 24: 115-124.
- Stebbins GL. Taxonomy and the evolution of genera, with special reference to the family Graminea. Evolution 1956. 10: 235-245.
- Sui X, Wang L, Xia X, Wang Z, He Z. Development of an allele-specific marker for Glu-B3 allele in common wheat and establishment of a multiplex PCR assay. Crop & Pasture Sci 2010. 61: 978-987.
- Tamás L, Shewry PR. Heterologous expression and protein engineering of wheat gluten proteins. J. Cereal Sci 2006. 43: 259-274.
- Takata K, Yamauchi H, Nishio Z, Kuwabara T. Breadmaking quality of a near-isogenic line with specific low molecular weight glutenin components. Breed. Sci 2001. 51: 143-146.
- Tao HP, Kasarda DD. Two-dimensional gel mapping and N-terminal sequencing of LMW-glutenin subunits. J. Exp. Bot 1989. 40: 1015-1020.
- Vasil V, Castillo AM, Fromm ME, Vasil Ik. Herbicide resistant fertile transgenic wheat plants obtained by microprojectile bombardment of regenerable embryogenic callus. BioTchnol 1992. 10: 667-674.
- Veraverbeke WS, Delcour JA. Wheat protein composition and properties of wheat glutenin in relation to breadmaking functionality. Crit. Rev. Food Sci. Nutri 2002. 42: 179-208.
- WallJS. The role of wheat proteins in determining baking quality. In: Laidman DL, Jones WRG. Recent advances in the biochemistry of cereals. (Eds). 1979. pp. 275-311.
- Wang LH, Li G, Pena RJ, Xia X, He Z. Development of STS markers and establishment of multiplex PCR for Glu-A3 alleles in common wheat. J. Cereal Sci 2010. 51: 305-312.
- Wang LH, Zhao XL, He ZH, Ma W, Appels R, Peña RJ, Xia XC. Characterization of low-molecular-weight glutenin subunit Glu-B3 genes and development of STS markers in common wheat (Triticum aestivum L.). Theor. Appl. Genet 2009. 118: 525-539.
- Weeks JT, Anderson OD, Blechl AE. Rapid production of multiple independent lines of fertile transgenic wheat (Triticum aestivum). Plant Physiol 1993. 102: 1077-1084.
- Weegels PL, van de Pijpekamp AM, Graveland A, Hamer RJ, Schofield JD. Depolymerisation and re-polymerisation of wheat glutenin during dough processing. I. Relationships between glutenin macropolymer content and quality parameters. J. Cereal Sci 1996. 23: 103-111.
- Wieser H. Chemistry of gluten proteins. Food Micro 2007. 24: 115-119.
- Zhao XL, Xia XC, He ZH, Gale KR, Lei ZS, Appels R, Ma W. Characterization of three low-molecularweight Glu-D3 subunit genes in common wheat. Theor. Appl. Genet 2006. 113: 1247-1259.
- Zhao XL, Xia XC, He ZH, Lei ZS, Appels R, Yang Y, Sun QX, Ma W. Novel DNA variations to characterize low molecular weight glutenin Glu-D3 genes and develop STS markers in common wheat. Theor. Appl. Genet 2007. 114: 451-460.
- Zhang W, Gianibelli MC, Rampling LR, Gale KR. Characterization and marker development for low molecular weight glutenin genes from Glu-A3 alleles of bread wheat (Triticum aestivum L.). Theor. Appl. Genet 2004. 108: 1409-1419.
- Zhang X, Jin H, Zhang Y, Liu D, Li G, Xia X, Zhonghu H, Zhang A. Composition and functional analysis of low-molecular-weight glutenin alleles with Aroona near-isogenic lines of bread wheat. BMC Plant Biology 2012. 12: 0-243.