적요
Genetically modified (GM) crops have been developed worldwide through the recombinant DNA technology and commercialized by various agricultural biotechnology companies. Commercialization of GM crops will be required the assessment of risk associated with the release of GM crops. The purpose of this research is a molecular characterization of introduced T-DNA in transgenic rice T4~T6 generation lines harboring a pepper MsrB2 gene under the control of stress inducible Rab21 promoter, as a part of biosafety evaluation for drought-tolerant transgenic rice (Agb0103). We identified the structure and sequence of transformation vector of T-DNA and analyzed insertion sites, flanking sequences, and generational stability of inserted T-DNA in transgenic rice lines. The transformation vector was consisted of right border, a drought-tolerant CaMsrB2 gene unit (Rab21 promoter:CaMsrB2:PinII terminator), a selectable marker herbicide resistance unit (CaMV 35S promoter:bar:Nos terminator), and left border in sequential order. Based on the adaptor-ligation PCR and whole genome sequence database, we confirmed that T-DNA was introduced 2 copies (head to head type) at the position of 2,471,957∼ 2,472,049 bp of chromosome No. 8. From the generational stability study, T-DNAs were stably inherited through the T4 to T6 generations, and also stable expression of bar gene from T-DNA was confirmed. It was also confirmed that the backbone DNA of transformation vector containing antibacterial gene was not present in Agb0103 rice genome. These results will be filed to biosafety assessment document of Agb0103.
서 언
유전자변형(Genetically modified, GM) 작물은 2013년에 1억 7,530만 헥타르가 재배되어 처음 상업적으로 재배되기 시작한 1996년보다 무려 100배 이상 증가하였고, 전 세계 27 개국에서 GM작물이 재배 승인된 것으로 농업생명공학 응용 을 위한 국제서비스(ISAAA)는 보고하고 있다(
James, 2013).
GM작물은 작물 생산에 필요한 노동력과 농기계, 농약 등의 사용 절감에 따른 경제적인 이익뿐만 아니라(
Owen, 2000),무경운 농법에 의한 토양 환경 보존 및 화석연료 절감과 급변 하는 기후 변화 등의 농업환경 변화에 대응이 가능케 한다 (
Brookes & barfoot, 2006). 하지만, GM 작물이 재배되기 시작한 1996년부터 재배 면적의 확대와 필요성과 함께, GM 작물 재배 지역의 환경 및 생태계 위해성과 식품 섭취에 따른 인체 영향 및 안전성에 대한 우려도 여전히 제기되고 있는 실 정이다.
GM 작물은
Toriyama 등(1988)에 의해서 최초 형질 전환 체 벼(
Oryza sativa)가 개발됨으로써 농업생명공학 분야에 큰 관심을 일으키며 연구가 시작되었으며(
Toriyama et al. 1988), 미국, 일본, 중국, 인도 등에서 활발하게 GM 벼 개발이 이루 어지고 있다. 2013년까지 우리나라에서 상업적으로 재배가 승인된 생명공학작물은 없으나, 제초제 저항성, 영양성분 강 화, 해충저항성, 가뭄저항성 등의 다양한 기능의 유전자들이 도입된 GM 벼의 개발이 집중적으로 이루어지고 있다(
Ha et al. 2010,
Jung et al. 2004,
Oh et al. 2010,
Shin et al. 2009). 이에 개발된 GM 작물이 독성, 알레르기와 영양 성분 평가 등에 관한 식품안전성 및 환경에 미치는 잠재적 위험성, 즉 도입 유전자들이 표적 및 비표적 생물체로 전이될 가능성 과 잡초화, 생태계 교란 등에 대한 환경위해성뿐만 아니라, GM 작물에 도입된 유전자의 발현 및 후대안정성 평가에 대 한 가이드라인 구축의 필요성이 대두되고 있다(
Oh et al. 2012). GM 작물들이 세대 진전에 따라 도입유전자의 발현정 지, 유전자 변이 등이 나타날 수 있어 GM 작물의 상업에 위 해서는 세대 진전에 따른 도입 유전자의 후대안정성 평가가 필수적이다(
Xia et al. 2009).
전 세계적으로 물 사용량 중에서 농업으로 인한 소비량이 가장 큰 비중을 차지하며, 개발도상국을 포함한 건조지역의 농업용수가 차지하는 비율은 전체 물 소비의 90% 이상을 차 지한다(
GLAAS 2012). 2012년 전 세계적인 가뭄으로 인해, 미국은 1,000여 개 이상 지역에서 재난선포를 한 바 있으며, 콩, 옥수수 등 곡창지대의 생산성 감소로 애그플레이션까지 유발시켰으며, 우리나라도 가뭄으로 인한 큰 피해를 입은 바 있다. 전 세계적으로 물은 점점 부족하게 될 것이며, 작물에 대한 농업 용수의 효율성이 중요한 문제로 제기되고 있다 (
FAO 2004,
UNESCO 2012). 이러한, 물 부족에 대비하여 물 자원의 관리와 농업지속성을 위한 체계적 접근과 더불어 생명공학 기술을 이용하여 내건성 작물 개발의 필요성이 더 욱 증대되고 있다. 최근, 몬산토에서 개발된 가뭄저항성 옥수 수는 상업화가 되었으며, 한국에서도 감자 유래
StMYB1 유 전자(
Im et al. 2012), 대장균 유래 Trehalose 생합성 유전자 (
Jang et al. 2003), 애기장대의
PUB22, 23 (
Cho et al. 2008) 등의 가뭄저항성 기능 검정이 보고되었다.
일미벼를 모본으로 생명공학기술을 이용하여 개발된 가뭄저 항성 벼는 스트레스 유도성 프로모터하에 고추 유래
CaMsrB2 유전자를 도입하여 개발되었으며, 유묘기와 성숙기에 강한 건 조저항성을 나타낸다. 고추에서 분리한
CaMsrB2 유전자는 메티오닌 잔기의 산화를 안정적인 환원상태로 전환시키는 기 능을 통하여 식물의 생물 및 비생물적 스트레스에 대한 저항 성을 부여하는 유전자이다. 가뭄 등의 원인에 의한 엽록체내 에서 산화스트레스 조건에서 메티오닌의 산화를 방지하고 미 토콘드리아, 엽록체, 그리고 퍼옥시좀에서 생산되는 활성화산 소(ROS)의 제거 기작을 통해 엽록체의 손상을 방지함으로서 가뭄 등 스트레스 저항성을 유도한다(
Kim et al. 2014,
Oh et al. 2010).
본 시험에서는 고추 유래 CaMsrB2 가 도입된 가뭄저항성 벼(Agb0103)의 도입 유전자들의 후대안정성 분석을 위해서 가뭄저항성벼(Agb0103)에 도입된 목적유전자(CaMsrB2), 개 시인자(Rab21 and 35S), 종결인자(NOS and PinII) 및 선발 마커(bar) 유전자들의 3세대에 대한 서던 분석과 목적유전자 발현 검정을 위한 RT-PCR 분석, 생육시기-부위별 PAT 발현 분석을 수행하였다. 본 시험을 통해 국내 개발 GM 작물의 안 전성 자료 생산뿐만 아니라, 3세대 이상의 복수 세대에 대한 GM 벼의 후대안정성 평가 및 안전성 평가 기준 설정을 위한 기초 자료로 활용하고자 하였다.
재료 및 방법
도입유전자의 인접서열 분석
가뭄저항성벼(Agb0103)의 게놈 DNA에 도입된 T-DNA 의 인접서열을 확인하고자
Ha 등(2010)의 방법에 따라 두 단 계로 실시하였다. 가뭄저항성벼(Agb0103)의 T4와 T6세대의 게놈 DNA를
Hae III로 절단하고, AD1와 AD2으로 구성된 adaptor를 접합하였다. 두 번째 단계로 adaptor에 대한 A1 (5’-TAATACGACTCACTATAG-3’)과 A2 (5’-GACTCACTATAGCAATTAAC- 3’)의 특이 프라이머를 제작하고, T-DNA Right border에 대해서 RB1 (5’-GCTTGCTGAGTGGCTCCTTC- 3’)과 RB2 (5’-CAAACGTAAAACGGCTTGTCC -3’)를 Left border에 대해서는 LB1 (5’-GTCAGGAGCTCGAATTCAGT- 3’)과 LB2 (5’-CATTAAAAACGTCCGCAATG- 3’)의 프라이머를 제작하여 단계별로 PCR을 수행하 였다. 이후 증폭된 PCR산물을 1% 한천 겔에 전기영동하고 Gel Extraction Kit (Qiagen, 28704)를 이용하여 정제 후, pGEM T-easy 벡터(Promega, Madison, USA)에 삽입하여 염기서열 정보를 확인하였다. 확인된 염기서열은 NCBI blast 분석을 이용하여 동일성 분석을 수행하였다.
가뭄저항성벼(Agb0103 T4, T5, T6)와 일미벼 식물체 시 료를 각 1 g씩 취하여, 막자 사발에서 액체질소와 함께 분말 화한 후 DNeasy Plant kit (Qiagen, Valencia, CA)를 이용하 여 게놈 DNA를 분리하였다. NanoDrop Spectrophotometer ND-1000 (NanoDrop Technologies Inc, Wilmington, DE) 을 이용하여 260/280 nm 값이 1.8~2.0 사이인 DNA 추출 액을 실험에 이용하였다. 가뭄저항성벼의 도입유전자 확인을 위해서 추출한 게놈 DNA 10
μg를 제한효소
BamH I과
Pme I 또는
BsrB I으로 처리하여 절단하고 1% 한천 겔에서 전기 영동후 변성화 과정을 수행하였다. 이 후 nylon membrane (Hybond-N+, Amersham, Uppsala, Sweden)에 겔의 DNA 를 전이시키고, membrane의 DNA 단편들을 UV-crosslink (1,200 × μJ/cm
2)로 고정한 후 혼성화 용액(0.5 M Na
2PO
4 pH 7.2, 1% BSA, 7% SDS, 1 mM EDTA, 10 mg/mL salmon sperm testicle DNA)에 1시간 동안 전 처리하였다. 혼성화는 Random primer DNA labeling kit (Takara Bio Inc., Shiga, Japan)를 이용하였고, 방사선인 α-
32P로 표지하 여, 65°C에서 16~18시간 동안 혼성화하였다. 유전자 운반체 (T-DNA) 내의 구성 요소들의 유전적 후대안정성 검정을 위 한 프로브는 특이적인 프라이머를 이용하여 목적 유전자 (
CaMsrB2), 프로모터(
35S,
Rab21), 종결인자(
NOS,
PinII) 및 선발마커 유전자(
bar)를 PCR로 증폭한 후 정제하여 사용 하였다(Table
1). Backbone DNA 유무 검정용 프로브는 항 생제인 spectinomycin의
aadA (SPM) 유전자 부위와 pSB11 5101-6100 부위를 PCR로 증폭한 후 elution 하여 사용하였 다(Table
2). 혼성화가 끝난 membrane은 세척액(1st solution, 2X SSC, 0.1% SDS; 2nd solution, 1X SSC, 0.1% SDS; 3rd solution, 0.2X SSC, 0.1% SDS)으로 세척한 후 Bioimaging analyzer (BAS-2000, Fuji Photo film, Tokyo, Japan) 로 분석하였다.
Table 1.Primers list used for the Drought-tolerant transgenic rice (Agb0103) Southern blot analysis.
Table 1.
|
Probe |
Primer |
Primer sequence |
Product Size (bp) |
|
CaMsrB2
|
CaMsrB2 Forward |
5'-TTGCGATGGCTACACCAGGCTC-3' |
348 |
|
CaMsrB2 Reverse |
5'-GGTTCGGGAACCCCTCACCCTT-3' |
|
bar
|
bar Forward |
5'-CTGCACCATCGTCAACCACT-3' |
505 |
|
bar Reverse |
5'-GGAAATTCGGGGTCATCAGA-3' |
|
Rab21
|
Rab21 Forward |
5'-TCTCCCGCCGCGTGCAATAC-3' |
391 |
|
Rab21 Reverse |
5'-CTAGCGTGGTCGCCGGCATC-3' |
|
Pin II
|
Pin II Forward |
5'-TGGGCATCAAAGTTGTGTGT-3' |
593 |
|
Pin II Reverse |
5'-TTGACAATCCATTCGTTTCT-3' |
|
NOS
|
NOS Forward |
5'-ATTGAATCCTGTTGCCGGTC-3' |
237 |
|
NOS Reverse |
5'-AGCTTGATATCGAATTCCCG-3' |
|
35S
|
35S Forward |
5'-ACGCAGCAGGTCTCATCAAG-3' |
554 |
|
35S Reverse |
5'-ATCGCAATGATGGCATTTGT-3' |
Table 2.Primers used for the backbone DNA detection PCR analysis.
Table 2.
|
Probe |
Primer |
Primer sequence |
Product Size (bp) |
|
pSB11 131-1300 |
131-1300 Forward |
5'-ATTACTCCGGCGGCCTTAGA-3' |
1,170 |
|
131-1300 Reverse |
5'-TCGGATAGACCTGCCTGTGC-3' |
|
pSB11(SPM) 1304-2092 |
1304-2092 Forward |
5'-CTTCGCGCCGCACCTCAACTG-3' |
789 |
|
1304-2092 Reverse |
5'-GTAACGCGGGCCGAAACAAACC-3' |
|
pSB11 2101-3400 |
2101-3400 Forward |
5'-CTGCACCATCGTCAACCACT-3' |
1,300 |
|
2101-3400 Reverse |
5'-GGAAATTCGGGGTCATCAGA-3' |
|
pSB11 3501-5100 |
3501-5100 Forward |
5'-GGGACCCGTTGTATCTGGTT-3' |
1,600 |
|
3501-5100 Reverse |
5'-TCGGATGGGGTTTTATAGGG-3' |
|
pSB11 5101-6100 |
5101-6100 Forward |
5'-TGGGCATCAAAGTTGTGTGT-3' |
1,000 |
|
5101-6100 Reverse |
5'-TTGACAATCCATTCGTTTCT-3' |
|
RAc1 |
RAc1 Forward |
5’-ATCACTGCCTTGCTCCTAGC-3’ |
350 |
|
RAc1 Reverse |
5’-GTACTCAGCCTTGGCAATCC-3’ |
Backbone DNA의 도입 유무 PCR 검정
가뭄저항성벼 Agb0103의 게놈 DNA에 유전자 운반체의 T-DNA 이외의 backbone DNA 삽입유무 확인을 위한 유전 자 운반체의 유전정보를 바탕으로 가뭄저항성벼의 backbone DNA (pSB11 vector) 확인용 프라이머를 제작하였다(Table
2). PCR 검정을 위하여 dNTP (10 mM) 4 μl, 10X PCR buffer 4 μl, 프라이머 각 20 μM, f-Taq DNA polymerase 1 unit (Solgent, Korea), 주형 DNA로 pSB11-RbMSR 10 ng 과 가뭄저항성벼 (T4, T5, T6) DNA 1 μg를 추가한 후 최종 반응 부피를 40 μl로 하였다. PCR 반응은 PTC-100 Thermal cycler (MJ Research, USA)를 이용하여 1 cycle (95°C, 5분), 35 cycle (95°C, 40초–60°C, 40초–72°C, 60초), 1 cycle (72°C, 5분)반응을 순차적으로 실시하였다. 증폭된 PCR산물 은 1% 한천 겔에서 전기 영동한 후 UV조사로 확인하였으며 Gel Extraction kit (Qiagen, 28704)를 이용하여 정제하고 pGEM T-easy 벡터 (Promega, Madison, USA)에 삽입하여 정확한 염기서열 정보를 확인하였다.
가뭄저항성벼 Agb0103의 T4, T5, T6와 일미벼 식물체 시 료를 각 1 g씩을 취하고, 막자 사발에서 액체질소와 함께 분 말화한 후 RNeasy Plant mini kit (Qiagen, CA, USA)을 이 용하여 RNA를 분리하였다. NanoDrop Spectrophotometer ND-1000 (NanoDrop Technologies, Inc, Wilmington, USA) 를 이용하여 260/230 nm 값이 2.0 이상인 RNA추출액을 실험 에 이용하였다. 추출한 total RNA 5 μg를 주형으로, RevertAid
TM First strand cDNA synthesis Kit (Fermentas, CA, USA) 이 용하여 총 볼륨 20 μl로 cDNA를 합성하였다. 식물 형질전환 용 운반체의 유전정보를 바탕으로 가뭄저항성벼의
CaMsrB2 와
bar 유전자의 발현 확인용 프라이머셑을 제작하였고 (Table
1), 벼 내재 유전자인 액틴을 검출하기 위하여 RAc1 F (5’-ATCACTGCCTTGCTCCTAGC-3’)과 RAc1 R (5’- GTACTCAGCCTTGGCAAT CC-3’)의 primer를 제작하여 cDNA 검정용으로 사용하였다. 합성된 일미벼와 가뭄저항성 벼(T4, T5, T6)의 cDNA를 5 μl씩, 양성대조군으로 유전자 운반체인 pSB11-RbMSR 10 ng를 주형DNA로, dNTP (10 mM) 4 μl, 10X PCR buffer 4μl, 프라이머 각 20 μM, f-Taq DNA polymerase 1 unit (Solgent, Korea)를 추가한 후 최종 반응 부피를 40 μl로 PCR 검정을 수행하였다. PCR 반응은 PTC-100 Thermal cycler (MJ Research, USA)를 이용하여 1 cycle (95°C, 5분), 35 cycle (95°C, 40초 - 55°C, 40초 - 72°C, 40초), 1 cycle (72°C, 5분) 반응을 순차적으로 실시하 였다. 증폭된 PCR산물은 1% agarose gel에서 전기 영동한 후, UV조사로 확인하였으며 Gel Extraction kit (Qiagen, 28704)를 이용하여 정제하고 pGEM T-easy 벡터(Promega, Madison, USA)에 삽입하여 정확한 염기서열 정보를 확인하 였다.
PAT (
bar) 유전자의 발현 확인을 위하여 Immu
NOStrip 검정 (lateral flow strip test)을 실시하였다. 가뭄저항성벼 (Agb0103 T4~T6)와 일미벼 식물체 시료를 추출액과 함께 마쇄하여 단백질을 추출한 후, PAT (
bar) 유전자의 발현을 Trait LL Test Strip (Strategic Diag
NOStics Inc, Newark, DE)를 이용 하여 Immu
NOStrip 검정을 수행하였다(
Oh et al. 2013). PAT 단백질의 농도를 정량하기 위하여 가뭄저항성벼 Agb0103 (T6)와 일미벼의 성장 시기별로 잎을 채취하여 시료들을 마 쇄한 후 PBST용액과 함께 균질화 한 후 저온처리(얼음에서 5분) 및 원심분리(5,000 g, 5분)하여 단백질을 분리, 추출한 후 PAT/
bar ELISA kits (EnviroLogix LibertyLink, Portland, USA)를 이용하여 ELISA분석을 실시하였다. 모든 시료들은 상온에서 2시간 반응한 후 ELISA reader (Multiskan EX, Thermo Scientific)를 이용하여 450 nm 파장에서 흡광도를 측정하였다(
Oh et al. 2013).
결과 및 고찰
도입 유전자의 도입위치 및 주변 염기서열 분석
본 시험에 공시한 가뭄저항성벼는 일미벼를 모본으로 엽록 체 내에서 메티오닌의 산화를 방지하고 활성산소의 제거 기 작을 통해 엽록체의 손상을 방지하여 가뭄 등 스트레스 저항 성을 유도하는 고추 유래
CaMsrB2 유전자와 스트레스 유도 성 프로모터(
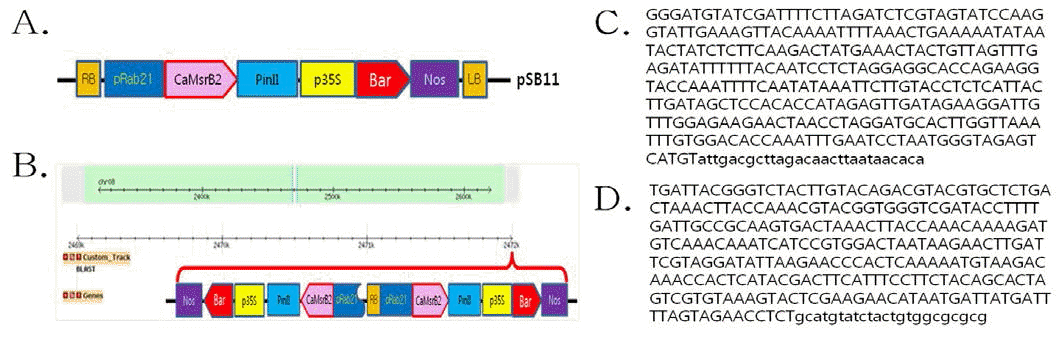
Rab21 promoter)를 부착하여 개발되었으며(Fig.
1A), 가뭄저항성벼 Agb0103 내의 도입유전자들의 전체 염기 서열 분석을 수행한 결과, 도입유전자 운반체내(pSB11- RbMsrB) 에 있는 주요 유전자 6 개에 대한 염색체내 삽입 및 완결성을 확인하였다. 그 결과, 도입유전자 두 카피가 Right Border (RB)를 중심으로 역방향으로 식물내에 도입됨을 확인하였다 (Fig.
1B,
Siddiqui et al. 2014). 가뭄저항성벼 Agb0103은 유묘 기, 성숙기에 강한 건조저항성을 나타낸다(
Kim et al. 2014). 벼의 유묘기와 성장기에서 가뭄저항성이 증진된 가뭄저항성 벼 복수 세대(Agb0103 T4~T6)에 대한 도입 유전자들의 유 전적 안정성 분석을 실시하였다. 우선, 도입유전자의 분자생 물학적 특성 분석은 가뭄저항성벼(Agb0103 T4, T6)의 T-DNA 삽입부위의 인접서열을
Ha 등 (2010)의 방법에 따라 단계적 으로 수행하였다. 먼저, 추출한 게놈 DNA를 제한효소
Hae III로 처리하여 1st PCR을 수행하였고, 이를 바탕으로 2nd PCR을 수행하여 한천 겔 전기영동으로 T-DNA의 Left border (LB) 부위의 증폭된 밴드를 확인하였고, 증폭된 첫번째 Left border (LB-1)와 두번째 Left border (LB-2)의 PCR 산물은 pGEM T-easy 벡터에 클로닝하여 염기서열을 분석하고, 이 들 염기서열에 대하여 NCBI blast 검색을 실시하여 확인한 인접서열을 Fig.
1C,
1D에 나타내었다. 검색 결과 운반체 내 의 T-DNA 이외의 염기는 확인되지 않아서, T-DNA만이 벼 의 게놈에 삽입되었음을 알 수 있었다. 또한 T-DNA의 인접 서열에 대한 게놈의 유사성 분석과 삽입위치를 확인한 결과, 유사성 있는 염기서열은 확인되지 않았으며, T-DNA의 삽입 부위는 도입유전자의 LB-1 및 LB-2의 인접염기서열을 Gene Bank (NCBI)에서
Oryza sativa (japonica cultivar-group) 게놈 데이터베이스에 근거하여 BLAST한 결과, LB-1 쪽의 염기서열은 벼 염색체 8번 중에서 2,470,655~2,470,960번, LB-2 쪽은 염색체 8번 중 2,471,052~2,471,327번 사이의 염기서열과 일치하였다. BLAST 염기서열 분석 결과를 바탕 으로 도입유전자의 삽입 위치를 확인한 결과, 벼 염색체 8번 중 2,470,960번과 2,471,052번 사이의 단일 부위에
CaMsB2 도입유전자가 삽입되어 있는 것을 확인하였다. 따라서 가뭄저 항성벼 Agb0103 T4와 T6의 주변 염기 서열 분석이 서로 일 치하는 결과를 나타냈으며, 가뭄저항성벼 Agb0103의 도입유 전자가 벼 8번 염색체 내에서 intergenic한 상태로 안정적으 로 유지되고 있음을 확인하였다(Table
3).
Fig. 1.
T-DNA junction sequences analysis.
(A) Schematic representation of the pSB11-RbMSR vector used for rice transformation. pSB11-RbMSR vector contained a recombinant
gene, CaMsrB2. The remaining construct components had the same configuration, consisting of the rice Responsive to ABA protein 21 promoter (Rab21), the 3’region of the potato proteinase inhibitor II gene (PinII) and the Bar expression cassette containing a 35S promoter/Bar coding region/3’region from the nopaline synthase gene (NOS). (B) Insertional site of T-DNA on chromosome in Agb0103 (C: left border-1; D: left border-2) for the Drought-tolerant transgenic rice (Agb0103) line flanking region. The genomic DNA was digested with Hea III, ligated to genome walking adapter to create different libraries, and used as template for PCR. Capital letters represents the flanking genomic sequences and lower case letters show the inner part of each border of T-DNA.
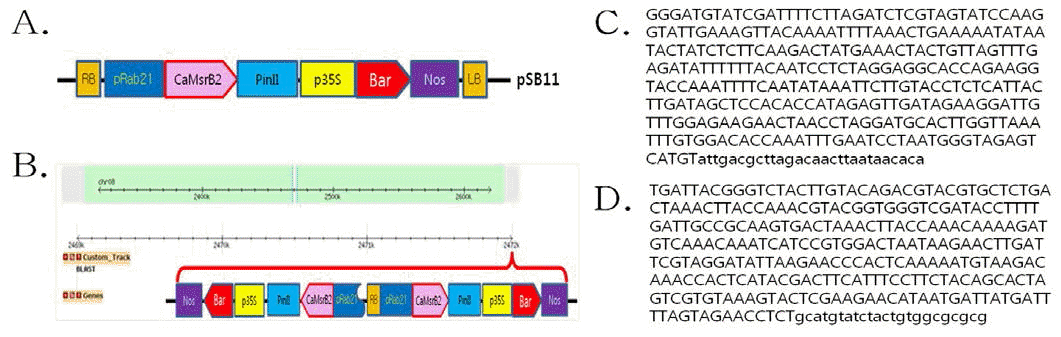
Table 3.Junction sequence analysis of T-DNA for the Drought-tolerant transgenic rice (Agb0103) line flanking region.
Table 3.
|
Pedigree |
Chromosome |
Junction sequence |
Junction gene & sequence |
|
|
Agb0103 |
8 |
2,470,655-2,470,960 2,471,052-2,471,327
|
Os08g0144000 2432861~2435744 Os08g0144100 2494083~2495012
|
도입 유전자 서던 분석
가뭄저항성벼의 도입 유전자(T-DNA)는 산화스트레스 저 항성 유전자인
CaMsrB2, 개시인자(
35S,
Rab21), 종결인자 (
NOS,
PinII), 및 선발마커(
bar) 유전자로 구성되어있다(Fig.
1A). 도입 유전자의 후대안정성 분석에 사용된 복수세대의 가뭄저항성벼 Agb0103 (T4~T6) 시료에서 도입 유전자들인 목적유전자(
CaMsrB2), 개시인자(
35S,
Rab21), 종결인자(
NOS,
PinII) 및 선발마커(
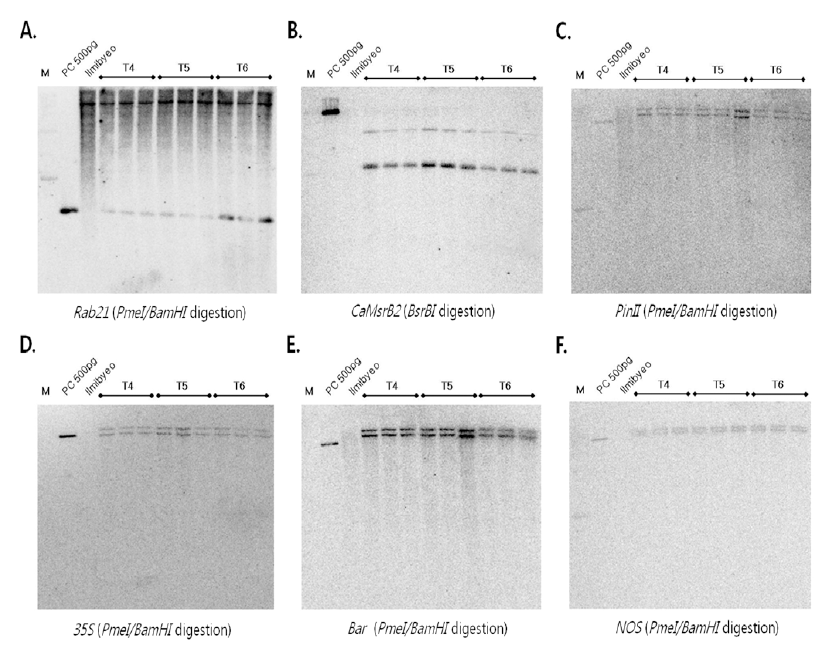
bar) 유전자에 대한 서던 분석을 수행하 였다. 가뭄저항성벼 Agb0103 T4~T6의 각 세대별 잎에서 게놈 DNA를 분리 추출한 후, 각 10 ug를
BamH I과
Pme I 또는
BsrB I 제한효소로 처리하여 단일 또는 삼중부위를 절 단하였고, 절단된 게놈 DNA를 1% 한천 겔에서 전기 영동한 후, Nylon membrane에 전이하였다. 양성대조군 (PC)으로는 pSB11-RbMSR 500 pg을 사용하였다. 가뭄저항성벼 세대별 (Agb0103 T4~T6, 제한효소
BamH I/
Pme I 절단)로
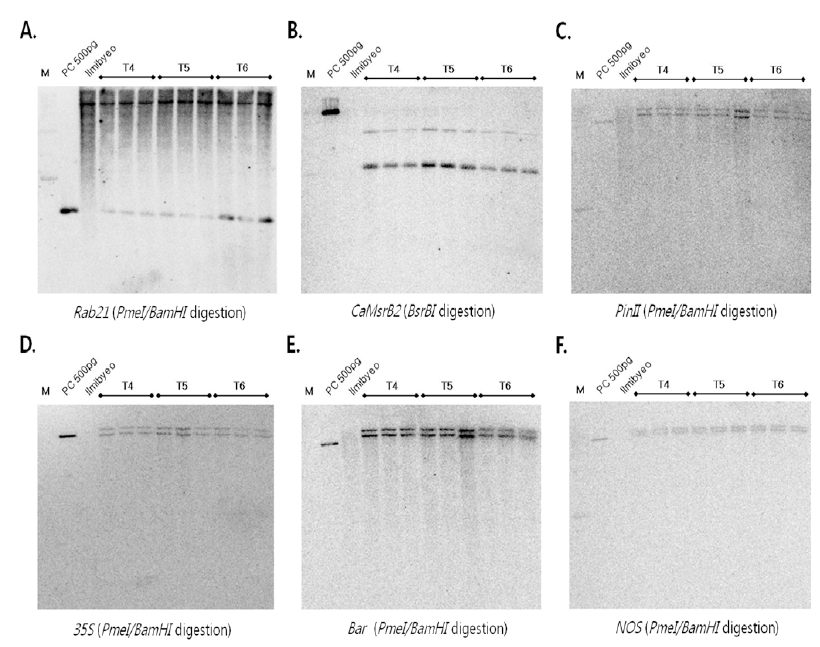
Rab21 프로모터를 프로브로 이용하여 서던 분석한 결과, 음성대조군 인 일미벼와 T4에서 T6세대까지의 가뭄저항성벼 Agb0103 에서 25kb 크기의 밴드가 검출되었고, 전체적으로 끌리는 양 상을 보였으며, 이는
Rab21 프로모터가 스트레스 유발에 의 해 발현되는 벼 내재 유전자이기 때문인 것으로 사료된다. 그 러나, 가뭄저항성벼 Agb0103에서만 2.6kb 크기의 밴드가 검 출되었고, 이 밴드는 가뭄저항성벼에 도입된
Rab21 프로모터 부분만 절단되어 검출된 것으로 확인되었다(Fig.
2A). 가뭄저 항성 유전자인
CaMsrB2 유전자(
BsrB I 절단)를 프로브로 이 용하여 서던 분석 결과, 복수세대의 가뭄저항성벼 Agb0103 의 복수세대(T4~T6세대)에서 모두 동일하게 6kb과 8kb의 두 개의 밴드가 확인되었고, 음성대조군인 일미벼에서는 검출 되지 않았다(Fig.
2B). 가뭄저항성벼의 각 세대별(Agb0103 T4~T6,
BamH I/
Pme I 절단)로
35S 프로모터 유전자를 이 용하여 서던 분석을 수행한 결과, 3세대 동안 안정적으로 동 일한 위치인 15kb과 20kb 크기의 두 개 밴드가 검출되었으 며, 일미벼에서는 검출되지 않음을 확인하였다. 이는 두 개의
35S 프로모터 유전자가 도입되었음을 확인할 수 있었다(Fig.
2D). 가뭄저항성벼 Agb0103 T4~T6세대 별로 게놈 DNA 를 제한효소
BamH I와
Pme I 로 절단하여 제초제 저항성 선 발 마커인
bar 유전자를 프로브로 사용하여 서던 분석을 수행한 결과, 일미벼에서는 검출되지 않았고, 가뭄저항성벼 Agb0103에 서만 15kb과 20kb 크기의 두 개의 밴드가 검출되어, 두 카피 의
bar 유전자가 도입되었음을 확인하였다(Fig.
2E). Agb0103 T4~T6 세대별로 도입유전자 터미네이터
PinII 및
NOS 터 미네이터 유전자의 서던 분석(각각
BamH I/
Pme I 절단) 결 과, 일미벼에서는 검출되지 않았으며, 가뭄저항성벼 Agb0103 T4, T5, T6에서만 15kb과 20kb 크기의 두 개의 밴드가 검출 되어, 모두 두 카피의
PinII 와
NOS 유전자가 도입되었음을 확인하였다(Fig.
2C,
2F). 이는 도입 유전자들인 목적유전자 (
CaMsrB2), 개시인자(
35S,
Rab21), 종결인자(
NOS,
PinII) 및 선발마커(
bar) 유전자들이 본 실험에 사용된 가뭄저항성벼 Agb0103에 두 카피로 도입되었고, 도입 유전자가 T4에서 T6 세대까지 안정적으로 발현됨을 확인할 수 있었다.
Fig. 2.
Genomic Southern blot analyses of Drought-tolerant transgenic rice (Agb0103).
Genomic DNAs from leaf tissues were digested with Pme I/BamH I, BsrB I and hybridized with probe (A) Rab21, (B) CaMsrB2, (C) PinII, (D) 35S, (E) Bar, and (F) Nos. The DNA molecular size markers are indicated on the left. PC: binary vector carrying CaMsrb2 recombinant genes as positive controls, Ilmibyeo: non-transgenic wild type rice as negative controls.
Backbone DNA의 도입 유무 PCR 검정
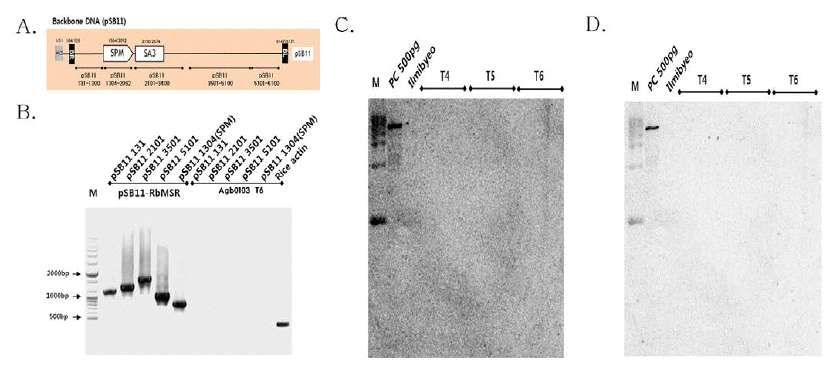
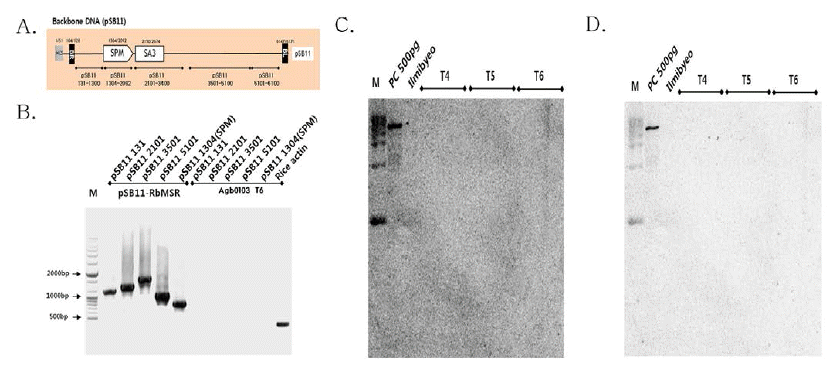
가뭄저항성벼 형질전환에 사용된 유전자 운반체의 backbone DNA인 pSB11 벡터에서 Fig.
3A와 같이 서던 분석용 프로 브 및 PCR 검정용으로 디자인하였으며, 항생제(spectinomycin) 내성 유전자(
aadA)를 포함한 총 5곳 부위를 프로브용 프라이 머를 제작하였고(Table
2), 가뭄저항성벼 Agb0103 T6의 게 놈 DNA를 Table
2의 프라이머 셑을 이용하여 PCR을 수행 하였다. PCR 결과, 가뭄저항성벼 형질전환에 사용된 pSB11- RbMSR (Backbone DNA와 T-DNA)에서는 각각의 부위에 따라 양성대조군(PC)인 backbone DNA에서만 밴드가 검출 되었고, 가뭄저항성벼 Agb0103 T6에서는 벼 내재 유전자인 액틴 유전자 밴드만 검출되어, Backbone DNA와
aadA (SPM) 가 도입되지 않았음을 확인하였다(Fig
3B). 가뭄저항성벼 Agb0103 T4~T6의 각 계통에 대한 항생제 내성 유전자(
aadA) 로 서던 분석 결과, 양성대조군(PC)인 pSB11-RbMSR에서만 밴드가 검출되었고, 가뭄저항성벼의 세대별(Agb0103 T4~ T6)에서는
aadA가 삽입되지 않음을 확인하였고(Fig.
3C), pSB11 벡터의 염기서열 5,101부터 6,100까지의 부분(pSB11 5101- 6100)을 프로브로 사용하여 서던 분석 결과, 양성대조군인 pSB11-RbMSR에서만 밴드가 검출되었고, 가뭄저항성벼의 세대별(Agb0103 T4~T6)에서는 pSB11 5101-6100가 삽입 되지 않음을 확인하였다(Fig.
3D).
Fig. 3.
Backbone DNA PCR and Southern blot analyses of Drought-tolerant transgenic rice (Agb0103).
(A) Schematic representation of the pSB11 vector used for PCR and Southern blot analyses. (B) Verification of Backbone DNA (pSB11) insert using PCR amplification of genomic DNA samples extracted from transgenic leaf tissues. (C) Genomic DNAs from leaf tissues were digested with Pme I/BamH I, and hybridized with probe aadA (SPM). (D) Genomic DNAs from leaf tissues were digested with Pme I/BamH I, and hybridized with probe pSB11 5101-6100. The DNA molecular size markers are indicated on the left. PC, PC means binary vector carrying CaMsrb2 recombinant genes as positive controls, respectively; Ilmibyeo, non-transgenic wild type (WT) rice as negative controls.
RT-PCR분석에 의한 목적유전자 발현 분석
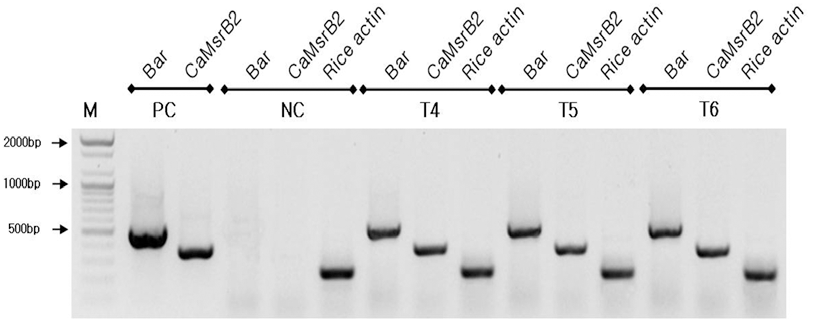
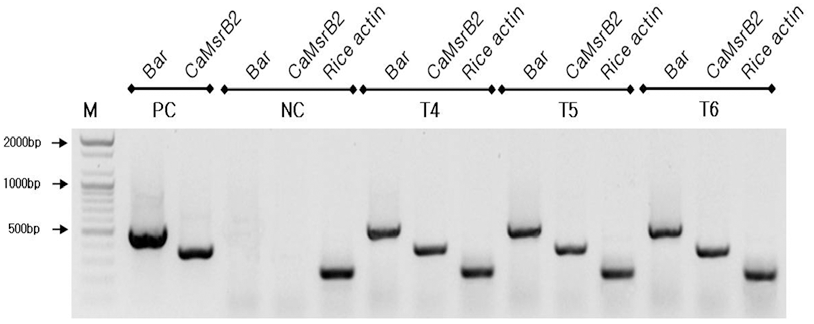
가뭄저항성벼 Agb0103의 세대간 안정적인 유전자의 발현 여부를 Reverse Transcriptase Polymerase Chain Reaction (RT-PCR) 분석을 실시하였다. 그 결과 연속되는 세대 간에 동일한 유전자 발현량을 나타냈으며(Fig.
4), 대조구인 일미 벼에서는 벼 내재 유전자인 액틴에서만 밴드가 검출되었고,
bar 와
CaMsrB2 유전자는 밴드가 검출되지 않았으며, 이는 가뭄저항성벼 Agb0103의 세대가 진전되어도 안정적으로 목 적유전자가 발현되고 있음을 확인하였다.
Fig. 4.
4. RT-PCR analyses of Drought-tolerant transgenic rice (Agb0103).
Two pairs of primers were to detect transcripts expressed in CaMsrB2 and bar gene for CaMsrB2 transgenic rice T4~T6 generations. The DNA molecular size markers are indicated on the left. PC: binary vector carrying CaMsrb2 recombinant genes as positive controls, NC: non-transgenic wild type (Ilmibyeo) rice as negative control.
도입유전자 복수세대의 단백질 발현 확인
가뭄저항성벼 Agb0103에서 선발마커로 사용된 제초제 저 항성인 PAT (
bar 유전자)단백질 발현을 검정하기 위하여 항 체를 이용한 lateral flow strip test (LFST) 분석을 실시하였 다. 가뭄저항성벼 Agb0103 T4~T6의 각 세대별로 파종한 후, 유묘기(파종 후 4주), 성장기(파종 후 8주), 출수기(파종 후 12주)와 등숙기(파종 후 16주)의 생육 시기별로 PAT 단백 질 확인용 항체가 표지되어 있는 immu
NOStrip을 이용하여 단 백질 발현을 분석한 결과, 세대별(T4~T6), 부위별(뿌리, 줄 기, 잎), 생육시기(유묘기, 성장기, 출수기, 성형기)별로 PAT 가 검출됨을 확인하였고, 대조군인 일미벼에서는 유묘기, 성 장기, 출수기, 성형기의 모든 부위(뿌리, 줄기, 잎)에서 PAT 가 검출되지 않았다(Fig.
5). 또한, 가뭄저항성벼 Agb0103에 서의 생육 시기(유묘기, 성장기, 출수기, 성형기)별로 PAT 단 백질 발현량을 정량하기 위해서 ELISA (enzyme-linked immu
NOSorbent assay)법을 이용하여 분석한 결과, 가뭄저항 성벼 Agb0103에서만 생육 시기(유묘기, 성장기, 출수기, 성 형기)별로 각각 14.04 ± 2.11
μg/g, 29.19 ± 3.95
μg/g 34.68 ± 1.83
μg/g 38.18 ± 9.71
μg/g 수준의 PAT 단백질이 발현되 었으며, 일미벼에서는 발현되지 않음을 확인하였다(Table
4). 본 실험에서는 가뭄저항성벼 Agb0103의 도입유전자에 대 한 후대안정성 평가를 수행하였다. 가뭄저항성벼 Agb0103 T4와 T6의 도입유전자 삽입 위치를 확인한 결과, 벼 8번 염 색체 중 2,470,960번과 2,471,052번 사이의 단일 부위에 intergenic한 상태로 Agb0103 도입유전자가 삽입되어 안정 적으로 유지되고 있음을 확인하였다. 또한, 도입된 T-DNA의 구성요소들이 3세대 이상의 복수세대에서 동일 위치내 2 카피 로 안정적으로 유지되고, 형질전환에 사용된 벡터의 backbone DNA는 도입되지 않았음을 확인하였다. 선발 마커로 도입된 PAT 단백질의 발현 분석을 통해 가뭄저항성벼 Agb0103 T4 에서 Agb0103 T6까지의 복수세대에서 생육시기별, 부위별로 안정적으로 발현되고 있음을 확인하였으며, 목적 유전자인
CaMsrB2와 선발 마커인
bar 유전자가 Agb0103 T4~T6 세 대에서 안정적으로 발현됨을 검증하였다.
Fig. 5.
Confirmation of bar gene expression for Drought-tolerant transgenic rice (Agb0103) by using immunostrip.
(A) Seedling stage, (B) Tillering stage, (C) Heading stage, (D) Ripening stage; Generation (Agb0103 T4~T6), Tissue(roots, stems,
leaves) type; Ilmibyeo, non-transgenic wild type rice as negative controls.

Table 4.PAT protein levels in Ilmibyeo and Drought-tolerant transgenic rice (Agb0103).
Table 4.
|
Sample |
PAT Concentration (μg/g) |
|
|
Seedling stage |
Tillering stage |
Heading stage |
Ripening stage |
|
|
Ilmibyeo |
0 (N.D.) |
0 (N.D.) |
0 (N.D.) |
0 (N.D.) |
|
Agb0103 |
14.04 ± 2.11z
|
29.19 ± 3.95 |
34.68 ± 1.83 |
38.18 ± 9.71 |
GM벼의 실용화를 위해서는 유전자변형생물체의 국가간 이동 등에 관한 법률에 따라 위해성평가와 심사를 거쳐야 하 지만 아직까지 경험이 많지 않아 평가항목에 대한 혼란이 개 발자 및 연구자들간에 존재한다. 따라서 향후 평가될 GM벼 들에 대한 후대안정성 평가에서는 본 실험에서 수행된 것처 럼 3세대 이상의 분자생물학적 특성 구명과 복수세대를 대상 으로 도입 유전자의 안정성, backbone DNA 삽입여부 및 목 적 유전자의 단백질 발현 평가가 이루어져야 할 필요가 있으 며, 향후, GM벼의 도입 유전자의 특성 검증 방법에 대한 안 전성 평가 기준과 데이타를 구축함으로써 보다 과학적이며 체계적인 GM작물의 후대 안정성 평가를 수행할 수 있을 것 으로 사료된다.
적 요
가뭄저항성벼의 복수세대에 대한 후대안정성을 서던 분석 과 PCR로 분석한 결과, 가뭄저항성벼의 Agb0103 T4~T6 세대에서는 도입된 모든 유전자들이 안정적으로 도입되어 있 으며, T-DNA 구성요소 이외의 backbone DNA는 가뭄저항성 벼에 삽입되지 않았음을 확인하였다. 목적 유전자인 CaMsrB2 와 제초제 저항성 선발 마커인 bar 유전자가 가뭄저항성벼 Agb0103의 T4~T6 세대에서 안정적으로 발현됨을 검증하 였다. 제초제 저항성 선발 마커로 도입된 PAT 단백질의 발현 분석 결과에서도 Agb0103 T4~T6의 3세대에서 생육시기별, 부위별로 안정적으로 발현됨을 입증하였다. 도입유전자의 삽 입 위치를 확인한 결과, 가뭄저항성벼 Agb0103의 도입유전 자가 벼 8번 염색체 내에서 intergenic한 상태로 안정적으로 유지되고 있음을 확인하였다. 이상의 분석 기법을 통해 복수 세대에서 가뭄저항성벼의 도입 유전자들이 안정적으로 유지 되고 목적 단백질들이 안정적으로 발현되고 있음을 확인하였다.
사 사
본 연구는 농촌진흥청 차세대 바이오그린21사업(과제번호: PJ009505012014)의 지원에 의해 이루어진 것임.
References
- Brookes G, Barfoot P. Global impact of biotech crops: Socio-economic and environmental effects in the first ten years of commercial use. AgBioForum 2006. 9: 139-151.
- Cho SK, Ryu MY, Song C, Kwak JM, Kim WT. Arabidopsis PUB22 and PUB23 are homologous U-Box E3 ubiquitin ligases that play combinatory roles in response to drought stress. Plant Cell 2008. 20: 1899-1914.
- FAO.Economic valuation of water resources in agriculture. 2004. FAO.
- GLAAS.UN-water global analysis and assessment of sanitation and drinking-water. 2012. WHO.
- Ha SH, Liang YS, Jung HR, Ahn MJ, Suh SC, Kweon SJ, Kim DH, Kim YM, Kim JK. Application of two bicistronic systems involving 2A and IRES sequences to the biosynthesis of carotenoids in rice endosperm. J. Plant Biotechnol 2010. 8: 928-938.
- Im JS, Cho KS, Cho JH, Park YE, Cheun CG, Kim HJ, Cho HM, Lee JN, Jin YI, Byun MO, Kim DY, Kim MJ. Growth, quality, and yield characteristics of transgenic potato (Solanum tuberosum L.) overexpressing StMyb1R-1 under water deficit. J. Plant Biotechnol 2012. 39: 154-162.
- James C. The global status of commercialized biotech/GM crops: 2013. ISAAA Briefs 45 2013.
- Jang IC, Oh SJ, Seo JS, Choi WB, Song SI, Kim CH, Kim YS, Seo HS, Choi YD, Nahm BH, Kim JK. Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose- 6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth. J. Plant Physiol 2003. 131: 516-24.
- Jung S, Lee Y, YANG K, Lee SB, Jang SM, Ha SB, BACK K. Dual targeting of Myxococcus Xanthus protoporphyrinogen oxidase into chloroplasts and mitochondria and high level oxyfluorfen resistance. Plant Cell Environ 2004. 27: 1436-1446.
- Kim JS, Park HM, Chae SH, Lee TH, Hwang DJ, Oh SD, Park JS, Song DG, Pan CH, Kim YH, Nahm BH, Kim YK. A pepper MSRB2 gene confers drought olerance in rice through the protection of chloroplasttargeted genes. Plos One 2014. 9: e90588.
- Oh SD, Park SY, Sohn SI, Kim JK, Park JS, Park SK, Cho HY, Ahn BO, Lee KJ. Evaluation and assessment of biosafety carotenoid-biofortified rice (PAC) : Progeny stability study for four generations. Korean J. Breed. Sci 2012. 44: 579-587.
- Oh SK, Baek KH, Seong ES, Joung YH, Choi GJ, Park JM, Cho HS, Kim EA, Lee S, Choi D. CaMsrB2, pepper methionine sulfoxide reductase B2, is a novel defense regulator against oxidative stress and pathogen attack. J. Plant Physiol 2010. 154: 245-261.
- Oh SD, Lee KJ, Park SK, Park HM, Lee SM, Ahn HI, Sohn SI, Cho HS, Ryu TH. Biosafety assessment for drought-tolerant transgenic rice (CaMsrB2-8) : Progeny stability study for three generations. Korean J. Breed. Sci 2013. 45: 109-118.
- Owen MDK. Current use of transgenic herbicide resistant soybean and corn in the USA. Crop Prot 2000. 19: 765-771.
- Shin KS, Lee SM, Lim SH, Woo HJ, Cho HS, Lee KR, Lee MC, Kweon SJ, Suh SC. Molecular biological characteristics and analysis using the specific markers of leaf folder-resistant GM rice. Plant Biotechnol. J 2009. 36: 115-123.
- Siddiqui ZH, Cho JI, Kwon TR, Ahn BO, Lee KS, Jeong MJ, Ryu TH, Lee SG, Park SC, Park SH. Physiological mechanism of drought tolerance in transgenic rice plants expressing capsicum annuum methionine sulfoxide reductase B2 (CaMsrB2) gene. Acta Physiol. Plant 2014. 36: 1143-1153.
- Toriyama K, Arimotoa Y, Uchimiyaa H, Hinata K. Transgenic rice plants after direct gene transfer into protoplasts. Nat. Biotechnol 1988. 6: 1072-1074.
- UNESCOManaging water under uncertainty and risk. UN Water Report 4 2012. 1: 22-69.
- Xia H, Lu BR, Su J, Chen R, Rong J, Song Z, Wang F. Normal expression of insect-resistant transgene in progeny of common wild rice crossed with genetically modified rice: its implication in ecological biosafety assessment. Theor. Appl. Genet 2009. 119: 635-644.