적요
The objective of this study was to identify quantitative trait loci (QTL) of spike length by association analysis with 94 doubled haploid wheat lines derived from Keumkang and Olgeuru. Days to heading date, culm length, spike length, and kernels per spike were evaluated in 2017 and 2018 in upland conditions. Culm length and spike length were biased short culm length and spike length (skewness=0.2 and 1.5, respectively). Kernel number per spike was biased low kernel number per spike (skewness=0.8). A genetic map was constructed with 170 microsatellite marker loci. One QTL was detected for spike length. The QTL on chromosome 4A, qSL-1 was detected by Xwmc283 and Xbarc327 explained 44.3% of the phenotypic variation. The QTL was applied to validate the relationship between genotypes of QTL and 29 Korean wheat cultivars grown for nine years under upland conditions. Korean wheat cultivars were classified into 5 types based on the combination of the two SSR markers. In Korean wheat cultivars, genotype b at Xbarc327, which was homozygous to Keumkang, had a shorter spike length (7.48 cm) than genotype c, which was different from the genotypes of Keumkang and Olgeuru, (8.45 cm). The two Korean wheat cultivars with genotypes ac at Xwmc283 and Xbarc327 had longer spike length (8.45 cm) than genotypes ba and bb (<7.42 cm).
서 언
밀은 전세계적으로 약 7억7천톤 정도가 생산되며, 전세계 인구가 소비하는 칼로리의 20%를 제공하는 중요한 식량원으로 국내에서도 연간 32.1 kg 소비되는 제2의 주식이다(
FAO 2019,
MAFRA 2019). 그러나 국내 밀 생산량은 년간 약 4만톤 생산되며, 곡물 자급도는 1.0% 미만으로 거의 전량을 수입에 의존하고 있는 상황에서, 정부는 밀산업육성법을 제정하였으며, 올해는 35년만에 국산 밀 수매 제도가 법에 명기되었다. 이러한 국산 밀 산업 체계의 변화를 지원하기 위해서는 무엇보다 국산 밀 생산의 안정성을 확보하는 것이 중요하다.
밀의 수량성은 주요 수량 구성 요소인, 단위면적당 수수, 일수립수와 천립중이 중요하며, 밀 생육 과정의 여러 농업 형질, 간장, 수장, 개화기와 재해 저항성등에도 영향을 받는다(
Fischer 2011,
Reynolds et al. 2012,
Valluru et al. 2014,
Nadolska-Orczyk et al. 2017). 밀 수량은 환경적으로 영향을 많이 받지만, 수량 관련 형질들 중에서 종실 무게 관련 특성은 다른 형질들에 비해 유전력이 높기 때문에, 관련 표지인자를 활용한 육종 초기 세대 선발에 적용 가능한 것으로 알려져 있다(
Hou et al. 2014,
Valluru et al. 2014,
Ma et al. 2015,
Wang et al. 2015,
2016,
Zhai et al. 2018). 이와 함께 수수와 일수립수 증가에 관련된 표지인자 탐색 및 활용을 통한 수량성 증대는 밀 육종 프로그램에서 매우 중요하기 때문에 관련 연구가 활발하다(
Reynolds et al. 2012,
Valluru et al. 2014,
Nadolska-Orczyk et al. 2017). 밀 수량은 많은 유전자가 상호 작용하는 양적 형질 위치(quantitative trait locus, QTL) 관련 표지인자가 이용되고 있으며, 육종 프로그램에서 표지인자 활용성을 높이기 위해 수량성 관련 QTLs, 전장 유전체 상관성 분석 연구(genome-wide association study, GWAS) 및 단일 염기 다형성(single nucleotide polymorphism, SNP) arrays의 이용에 대한 연구가 최근에 활발하게 진행되고 있다(
Bernardo 2008,
Valluru et al. 2014,
Lozada et al. 2017,
You et al. 2018).
국내 밀 품종 개발은 국내 이모작 재배에 적합한 조숙이면서 재해저항성을 지닌 수량성이 높고 가공 용도에 맞는 고품질 품종을 목표로 현재까지 약 50여 품종을 육성하였다. 이러한 성과로 이모작 재배 가능하고 다양한 가공 적성을 지닌 우수한 품종이 농가에 보급되어 재배되고 있지만, 농업인 및 생산 단체는 농가 소득 증대를 위한 국내 밀 품종의 수량성 향상을 요구하고 있다. 이러한 상황에서 국내 밀 육종프로그램은 최근 육성된 긴 이삭의 태중밀 육성 집단과 금강과 올그루를 교배본으로 만들어진 반수체 계통을 이용하여 수량성 관련 QTLs을 탐색하였다(
Lee et al. 2014,
Son et al. 2015,
Cho et al. 2018,
Kang et al. 2019). 본 연구에서는 금강과 올그루 유래 반수체 계통에서 추가적으로 탐색된 이삭 길이 관련 QTL의 국내 밀 품종의 적용을 통하여 국내 밀 육종 프로그램의 수량성 관련 선발 효율 증진을 위한 기초 자료를 제공하고자 한다.
재료 및 방법
공시재료
백립계 조숙품종인 금강과 추형의 이삭을 가진 올그루를 모⋅부본으로 국제밀옥수수 연구소(International Maize and Wheat Improvement Center, CIMMYT)에서
Inagaki & Mujeeb-Kazi (1995) 방법으로 금강과 올그루를 교배하여 얻은 F
1 개체에서 감수분열이 끝난 화분을 이용하여 약배양을 수행 후, 염색체 배가(chromosome doubling)하여 배가 반수체(doubled haploid, DH) 122개 생산하였으며, 이 계통을 국립식량과학원에서 증식하여 보관하였다(
Lee et al. 2014). 이들 계통 중에 모⋅부본을 포함한 무작위로 96계통을 선발하여 전북대학교 시험포장에서 전작조건(0.25 m×2 m×1열, 3반복)으로 2017년과 2018년 재배하였다. 연관분석을 통하여 확인된 QTLs에 상응하는 SSR (simple sequence repeat) 표지인자(marker)의 유효성을 확인하기 위하여 국내 29개 품종도 같은 조건으로 2010년부터 2018년까지 재배하였으며, 시비 및 재배는 농촌진흥청 표준 재배법(
RDA 2012)에 따라 수행하였다.
농업 형질 조사는 농촌진흥청 농업 과학 기술 조사 분석 기준(
RDA 2012)에 준하여 실시하였다. 출수기는 전체 단위 면적당 이삭 수 중에 40%가 출수한 날로 하였으며, 간장은 지면에서 이삭 목까지의 길이를 측정하였으며, 수장은 이삭 목에서 이삭 끝까지의 길이를 측정하였고, 일수립수는 생육이 균일한 장소에서 무작위로 20수를 채취하여 탈곡 후 조사하였다.
공시 재료를 전북대학교 온실에 파종하여 3주간 생육시킨 후, 엽을 채취하여 액체질소를 이용하여 분쇄한 뒤 -70℃에 보관하였다. 분쇄한 잎 100 mg으로부터 Genomic DNA분리시약(Solgent, 한국)를 이용하여 genomic DNA를 추출하였다. 추출된 gDNA는 Biodrop (Biodrop Ltd, 영국)를 통해 정량 및 정성분석을 한 후, 20 ng/uL로 희석하여 PCR분석에 사용하였다.
총 1,482개의 SSR 표지인자를 이용하여 다형성을 보이는 표지인자 192개를 선발하였다. SSR 표지인자를 사용하여 분리비 검정을 확인하였으며, 분리비 검정의 정확도를 높이기 위하여 SSR 표지인자에 상응하지 않는 결과(missing data)가 10%를 넘는 SSR 표지인자는 제외하여, 최종적으로 170개(11.47%)의 SSR 표지인자를 이용하였다. PCR조건은 touch-down 방법에 준하여 94℃에서 5분간 초기 변성 후 1차 반복에서 94℃에서 1분간 변성하고 66℃에서 62℃까지 1회당 1℃씩 감소하면서 1분간 결합, 그리고 94℃에서 1분간 확장을 5회 반복하고, 2차 반복에서94℃에서 1분간 변성하고 62℃에서 1분간 결합, 그리고 94℃에서 1분간 확장을 35회 반복하였다. PCR산물은 6% polyacrylamide gel 전기영동으로 확인하였으며, 전기영동 조건은 100 voltage에서 30분간 pre-running 후 120 voltage에서 약 50분간 전기영동하였다.
유전 지도 작성 및 QTL 분석
연관관계 분석을 위하여 DH 94계통을 이용하였다. 농업 형질 조사 결과를 바탕으로 한 표현형 분석(phenotypic analysis)과 SSR 표지인자를 이용한 유전형 분석(genotypic analysis)을 수행하여 이들 간에 연관관계(association analysis)를 분석하였다. 표현형 분석에 대한 수치화는 각 농업 형질 측정값의 평균값을 사용하였으며, 유전형 분석은 유전 분석 집단의 각 개체에 대하여 SSR 표지인자에 상응하는 모본과 부본의 유전형(homozygous type, A or B type)과 이형(heterozygous type, C type)을 판별하였으며, 유전자형의 분리비 이상(segregation distortion)을 검정하기 위하여 히트맵(heat map)을 응용하여 유전자형의 전반적인 분포 양상을 확인하였고 적합성 검정(
x2-test)을 통계프로그램 R (version 3.6.1,
https://www.r-project.org)을 이용하여 수행하였다. 연관 분석 결과를 바탕으로 MapMaker/EXP version 3.0b를 이용하여 유전 지도를 작성하였으며, QTL IciMapping version 4.0을 이용하였다(Wang et al. 2014). 각 형질 별 통계적인 유의성을 확인하기 위하여 순열검정법(permutation test,
n=1,000)을 수행한 결과, LOD 임계값(logarithm of odds threshold)은 3.05으로 계산되었다.
농업형질에 대한 표현형 분석에 사용된 모든 자료는 최소 3회 이상 반복 수행하여 평균값을 사용하였으며, 기본적인 기술통계는 통계프로그램인 R (The R Project for Statistical Computing, R version 3.4.4,
https://www.r-project.org)의 기둥그래프(histogram) 함수인 hist와 평균(mean), 최대값(maximum), 최소값(minimum), 표준편차(standard deviation), 분산(variance), 왜도(skewness), 및 첨도(kurtosis)의 각 함수인 mean, max, min, sd, skewness, 및 kurtosis를 이용하였다. 또한, SAS (statistical analysis system, SAS Institute, Cary, NC) 프로그램을 이용하여 QTL에 상응하는 표지인자를 이용한 국내 밀 품종에의 수장에 대한 유효성 검정 결과를 비교하게 위하여
t-test 및 분산분석(analysis of variance, ANOVA)를 수행하였다.
결과 및 고찰
표현형 및 유전 분석
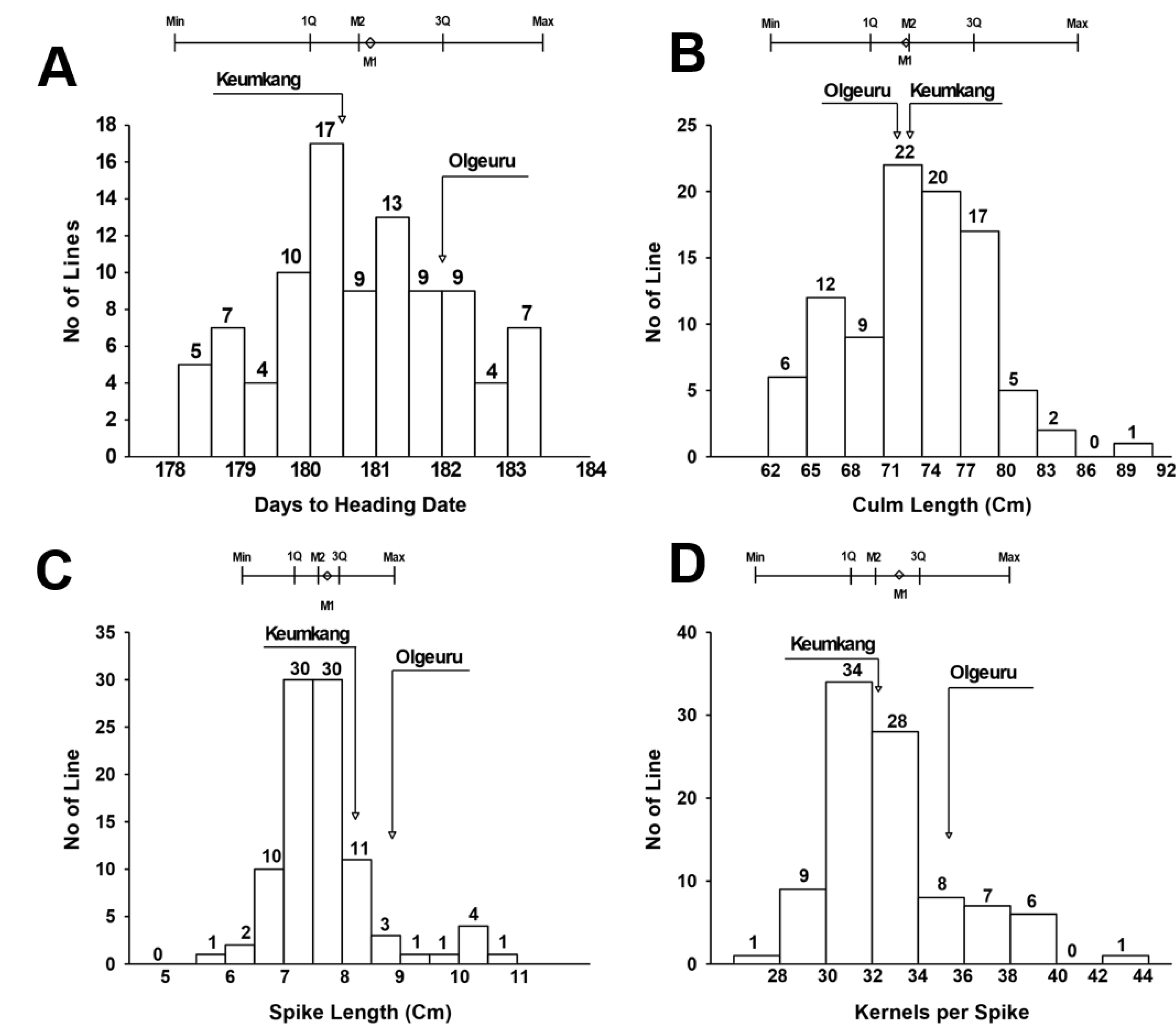
유전분석 집단(DH mapping population)의 표현형 분석을 위하여 2년 동안 전작조건에서 DH 94 계통에 대하여 출수일수, 간장, 수장, 및 일수립수를 조사하였다(
Table 1,
Fig. 1). 국내 밀 재배환경에서 숙기가 늦어져 장마철과 겹치면 수발아 뿐만 아니라 수량 및 품질 저하의 우려가 있기 때문에 밀 육종 프로그램에서 숙기 단축은 가장 중요한 목표였다. 숙기와 밀접한 관련이 있는 출수일수는 유전분석 집단에서 178일에서 183일 사이로 나타났으며, 평균 출수일수는 금강보다는 1일 그리고 올그루보다 2일 적은 180일로 확인하였다. 평균 출수일수를 기준으로 유전분석 집단의 분산은 정규분포를 나타내었다(skewness=0.0). 양친 인 금강과 올그루의 출수일수가 큰 차이가 없기 때문에 출수일수를 조절하는 유전자의 유전력은 낮은 침투율(penetrance)로 표현형에 큰 변화의 폭이 나타내지 않은 것으로 생각한다. 하지만 양친의 출수일수보다 빠른 출수일수를 보인 초월우성 계통들은 중요한 유전적 자원으로써 가치가 있다고 할 수 있다(
Fig. 1).
유전분석 집단의 간장과 수장의 범위는 각각 62.5-91.6 cm와 5.7-10.5 cm였으며, 평균은 각각 73.7 cm와 7.7 cm로 간장은 금강(72 cm)과 비슷하였고, 수장은 금강(8.5 cm)과 올그루(8.6 cm)보다 짧았다. 간장의 분산은 출수일수, 수장 및 일수립수 보다 변이 폭은 컸으며, 정규분포를 보였고, 수장은 평균을 기준으로 다소 짧은 수장쪽으로 편향된 양상(skewness=0.7)인 정적 비대칭(positive skewness)으로 나타났다(
Table 1). 일수립수는 27-43개였고, 평균 일수립수는 33개로 올그루(35개)보다는 적었고 금강(32개)과 유사하였으며, 일수립수의 분산은 간장과 유사하게 큰 변이 폭과 정규분포를 보였다.
SSR 마커 170개를 사용하여 분리비(segregation ratio)를 확인한 결과(
Fig. 2), 본 연구의 유전 분석 집단은 반수체 집단이기 때문에 동형 접합체이고 모⋅부본에 의한 분리비는 올그루(A type):금강(B type)=49.9:49.9였으며 이형(hetero)의 수는 0.2%로 거의 존재하지 않았다. 다형성을 보이는 170개의 SSR 마커 중 11개의 SSR 마커에 의하여 모⋅부본의 분리비 양상은 A type:B type=1:1로 나타났다. 이형 계통의 발현될 확률은 0.2%이지만, 170개의 SSR 마커 중, 48개 SSR 마커에 의하여 이형이 최소 2개에서 최대 8개까지 나타났다. 이중
Xgwm156에서는 특이적으로 28개가 이형으로 나타났다. 분리비 검정 결과, A type 유전자형을 갖는 개체의 수가 B type의 유전자형을 갖는 개체수보다 많은 분리비 이상(segregation distortion)을 나타내는 SSR 마커는 35개였다. 대부분의 SSR 마커에 의한 분리비 이상은 분리비 기대치와 고도로 유의한 수준에서 차이를 보였다. 올그루의 유전자형과 금강의 유전자형의 분리비 차이가 50% 이상인 SSR 마커는
Xgwm3,
Xbarc1과
Xgdm67이었다. B type 유전자형을 갖는 개체의 수가 A type의 유전자형을 갖는 개체수보다 많은 분리비 이상을 나타내는 SSR 마커는 40개였으며, 모부본의 유전자형 분리비 차이가 50% 이상인 SSR 마커는
Xbarc327, Xgwm471과
Xmag2152였다. 현저한 분리비 이상을 나타내는 6개의 SSR 마커를 제외한 12개 SSR 마커는 모⋅부본의 유전자형 분리비 차이가 약 20%에서 50% 미만이었다.
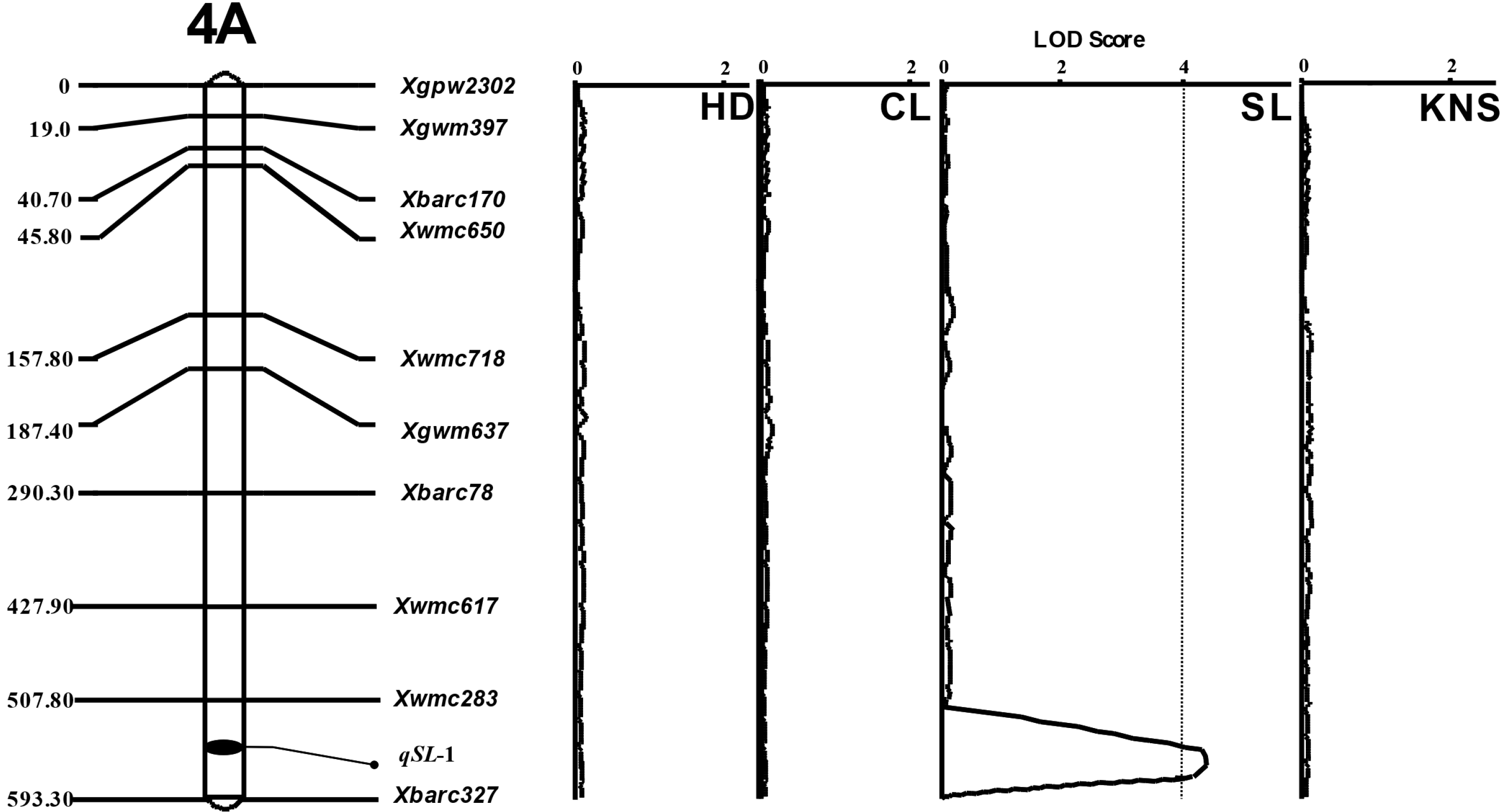
연관 분석을 수행하여 유전 지도를 작성하였으며, 출수일수, 간장, 수장과 일수립수에 대한 QTLs를 확인한 결과, 수장을 제외한 다른 형질은 관여하는 대립유전자가 표현형의 변화를 조절하는 유전력이 높지 않아 QTLs의 LOD 임계값을 넘지 못하여 유의성이 인정되지 않았다(
Fig. 3). 수장 관련 1개의 QTL이 4A (
qSL-1) 염색체에 위치하는 것으로 나타났다(
Table 2). 4A 염색체에 위치한
qSL-1 (LOD: 4.06)의 측면 마커(flanking marker)는
Xwmc283과
Xbarc327로, 전체 표현형 분산에 대한 QTL 기여도(phenotypic variation explained)는 44.3%로 나타났다.
qSL-1는 수장에 대한 전체 표현형 중 44.3%를 설명하는 주동적 대립 형질로 보이며, 상가적 효과(additive effect)는 1.16으로 금강의 대립 인자가 수장의 감소에 영향을 주었다.
기존에 수량과 관련하여 보고된 QTLs는 ITMI (International Triticeae Mapping Initiative) 집단에서 1A, 2A, 2B, 3B, 6D와 7A 염색체에 위치했고, 노르웨이의 반수체 집단에서는 수량 관련 QTLs는 게놈 전체에 고루 분포를 했으며, 17개 군집으로 나뉘었고, 이들 중 7개 군집의 영향이 컸으며, 7A과 7B염색체에 위치한 QTLs이 수량에 미치는 영향이 강했으며,
Rht1, Vrn-A1과
Vrn-D1이 위치한 4B, 5A와 5D염색체의 QTLs이 수량에 영향을 미쳤다(
Börner et al. 2002,
Li et al. 2002,
Jantasuriyarat et al. 2004,
McCartney et al. 2005,
Quarrie et al. 2005,
2006,
Kumar et al. 2007). 이러한 연구결과로 수량은 단일의 QTL에 의하여 설명되는 것이 아니라는 것을 알 수 있다. 또한, 기존의 연구에서 보고된 QTLs과 이 연구에서 확인된
qSL-1과는 염색체 위치와 기여도가 다른 것을 확인하였다.
중국 품종과 미국 적립계 연질밀과 교배로 만들어진 132계통(F
12)에서는 1A, 1B, 2B, 2D, 3B, 4A, 4B, 5A, 5B, 6A, 6B, 7A과 7D에서 10개의 QTLs가 보고되었고, 인도 유전 집단에서는 1D, 2D, 3B, 4A, 4D와 7A염색체에 위치했으며, 중국 유전 집단과 반수체 집단에서는 1A, 1B, 2A, 2B, 2D, 3A, 3B, 3D, 4A, 4D, 5A, 5B, 6D과 7D에 위치하였으며, 등숙과 종실 무게와 관련된 QTLs이 다면발현 효과를 나타내었다(
Marza et al. 2006,
Kumar et al. 2007,
Wang et al. 2009). 캐나다 반수체 집단에서는 53개 QTLs이 12개 염색체에 위치했고 1A, 2D, 3B과 5A 염색체에 위치한 5개 QTLs이 수량에 많은 영향을 주는 것으로 나타났는데, 캐나다 유전 집단에서도 수량 관련 QTLs가 5A, 7A와 7B 염색체에 위치했으며, 미국 빵용밀 반수체 집단에서는 6A 염색체에 위치하였고, 미국 연질밀에서는 1A, 2D, 3B와 4B 염색체에 위치 했으며, 일본 반수체 집단에서도 6번 염색체에 위치하였지만, 국내 반수체 집단에서는 2A (
Xgpw2206), 3A (
Xgwm247)와 3B (
Xwmc471)에 위치하였다(
Huang et al. 2006,
Cuthbert et al. 2008,
Hai et al. 2008,
Heidari et al. 2011,
Lee et al. 2014,
Simmonds et al. 2014,
Lozada et al. 2017).
국내 반수체 집단에서 수량과 관련 있는
Xgwm247은 이란 유전집단에서도 수량 변이를 설명하는 QTLs이었으며, 이들 집단에서는 이외에도 23 QTLs가 10개 염색체에 위치하였으며, 멕시코 봄밀 집단에서도 6A를 포함하여 2A, 3B, 4A와 4B 염색체에 수량 관련 SNP가 위치하였다(
Azadi et al. 2015,
Sukumaran et al. 2018). 반수체 집단에서는 17개의 QTLs가 수량과 엽록소 함량과 상관이 있었으며, 2D, 4A와 5D염색체의 QTLs은 수량과 엽록소 함량을 동시에 조절하는 것으로 나타났고, 엽록소 함량 및 PS II의 효율성 관련된 QTLs이 중국의 재조합 집단에서도 17개의 QTLs이 1A, 1B, 1D, 2D, 3D, 4A, 4B, 4D, 5A, 5D, 6D, 7A와 7D에 위치하였으며, 미국 연질밀 집단에서도 잎색에 관련된 QTLs이 2B 염색체에 위치하였다(
Zhang et al. 2009a,
2009b,
2010b,
Carter et al. 2011,
Parry et al. 2011). 미국 연질밀 집단에서도 수량은 숙기와 간장에 관련된 QTLs이 밀접하게 연관되어 있었으며, 병저항성이나 파성과 광주기성 등도 영향을 미치기 때문에 육종프로그램에서 농업 형질 선발에 유용한 마커 개발이 중요하다(
Carter et al. 2011). 최근에는 중국 품종에 대한 수량 관련 SNP 탐색을 통하여 12개의 SNP가 1A, 1B, 2A, 2B, 2D, 3A, 3B, 3D, 5A와 5B 염색체에 위치했으며, 1A (
AX_110387060,
AX_110418502), 1B (
AX_110508372,
AX_109820171), 2D (
AX_109941480), 3B (
AX_109881378)와 3D (
AX_95257733)에 위치한 SNP는 환경 변이에도 비교적 안정적인 것으로 나타났으며, 미국 적립계 연질밀 집단에서는 6A, 6B, 7A와 7D 염색체에 유용한 SNP가 위치하였다(
Li et al. 2019,
Ward et al. 2019). 60여개 논문을 분석한 meta-QTL 분석에서, 총 55 MQTLs을 찾았고 이중에서 1A, 1B, 2A, 2D, 3B, 4A, 4B, 4D과 5A 염색체에 위치한 12개 QTLs가 유효성이 높았고, 이들은
Rht 과
Vrn에 관련되었다(
Zhang et al. 2010a).
이삭 길이의 경우, 중국 품종과 CIMMYT 집단에서는 관련 QTLs이 주로 2A 염색체에 위치했으며, ITMI 집단에서는 1A, 1B, 2A, 2B, 2D, 4A, 6A와 7B 염색체에, 인도 유전 집단에서는 2A와 2D 염색체에, 일본 반수체 집단에서도 2D 염색체에, 중국 유전 자원과 유전 집단에서는 1A, 2B, 2D, 3B, 4A, 4B, 5A, 5B, 6B과 6D염색체에 위치하였고, 중국 품종과 미국 적립계 연질밀 교배로 만들어진 132계통(F
12)에서는 1A, 1B, 2B, 3B, 4B, 5B, 7A와 7B 염색체에 위치하였다(
Li et al. 2002,
Marza et al. 2006,
Crossa et al. 2007,
Kumar et al. 2007,
Ma et al. 2007,
Yao et al. 2009,
Liu et al. 2010,
Deng et al. 2011,
Heidari et al. 2011,
Cui et al. 2012,
Wu et al. 2014). 국내 반수체 집단에서도 2A, 2D, 3A, 5A와 6A 염색체에 위치한 이삭 길이 관련 QTLs는 10.7-12.6%의 이삭 길이에 대한 표현형을 설명하였으며, 국내 장수형 집단에서도 2A와 5B 염색체에서 이삭 길이에 관련된 QTLs 보고되었으며, 이 두 집단에서의 이삭 길이에 대한 QTLs의 위치는 비슷하였다(
Lee et al. 2014,
Kang et al. 2019). 국내 유전분석 집단의 QTLs는 ITMI 집단의 2A 염색체에서 보고된 QTLs와는 거리상 차이가 있었지만, 중국 유전자원의 2A 염색체에서 보고된 7개의 QTLs 중
Xgwm294 (76.4 cM)는 국내 반수체 집단과 장수형 집단에서 보고된
Xgpw2206 (76.5 cM)와
Xcfd5 (70.0 cM)와 비슷한 위치였다(
Kumar et al. 2007,
Yao et al. 2009,
Lee et al. 2014,
Kang et al. 2019). 하지만, 이 연구에서 수장과 관련된
qSL-1이 확인된 집단은 기존의 국내 반수체 집단과 동일한 모부본에서 파생된 다른 세대의 개체임에도 불구하고 기존에 보고된 수장과 관련된 QTLs과 다른 염색체 위치에 있는 것을 확인하였다. 이러한 이유는 동일한 모부본에서 파생되었지만 집단의 세대의 차이에 따른 표현형에 차이에 의한 것과 이 실험에서 사용된 SSR에 대한 정보는 일본 밀 유전자 지도에서 확인된 밀 SSR을 사용하였는데 국내 집단에서 SSR표지인자에 대한 다형성의 빈도가 11.5%로 매우 낮게 나온 것으로 미루어 보았을 때 SNP와는 다르게 SSR의 한계에 의한 것으로 생각한다. 따라서 표지 인자의 수와 표지 인자에 대한 유전적 정확도를 높이기 위하여 DNA chip을 이용한 고해상도(high resolution mapping)이 절대적으로 필요하다.
미국 연질밀 집단에서는 이삭 길이를 포함한 이삭 특성에 관련된 109개 SNP가 1A, 1D, 2B, 3B, 5B와 5D 염색체에서 보고되었으며, 유럽 품종에서는 2A, 2B, 2D, 3B와 6B 염색체에 위치하는 SNP 마커가 보고되었고, 중국 품종에서 57개 SNP가 1B, 2B, 2A, 2B, 2D, 3B, 6A와 6D 염색체에서 발견되었고, 중국 유전 집단에서는 40개 SNP가 1A, 1B, 2A, 2B, 2D, 3B, 3D, 4A, 4B, 5A, 5B, 6A, 6B, 6D, 7A, 7B와 7D 염색체에서 나타났는데, 2A 염색체에서 발견된 5개 SNP 위치가 78.0-78.3 cM으로 국내 품종에서 발견된 QTLs는와도 비슷한 위치로 보인다(
Sun et al. 2017,
Zhou et al. 2017,
Liu et al. 2018,
Würschum et al. 2018,
Li et al. 2019). 본 연구 결과와 마찬가지로 ITMI 집단과 중국 유전 자원에서도 4A 염색체에서 이삭 길이 관련 QTLs가 보고 되었는데, ITMI 집단에서 보고된
Xfba231 (93.5 cM)-
Xcdo545 (101.6 cM)는
Xgwm637 (93.9 cM)-
Xgwm160 (128.5 cM)과 비슷한 위치로 나타났지만, 중국 집단의 4A 염색체의 SNP는 염색체 말단 부위인 136.2-144.1 cM에서 6개의 SNP가 발견되었고(
wsnp_Ex_c1563_2987002 (136.2 cM), wsnp_Ex_c28092_37240192 (140.5 cM),
wsnp_Ex_rep_c67779_66463916 (141.4 cM),
wsnp_Ex_c2266_4247520 (141.8 cM),
wsnp_Ex_c12_21212 (143.0 cM),
wsnp_Ex_c3463_6348659 (144.1 cM), 이들과는 위치가 달랐으며, 다른 4A 염색체에서 보고된
IWB33160 (1.2 cM),
QSL.caas-4AS (57.6 cM),
QSL.caas-4AL.1 (146.0 cM),
QSL.caas-4AL.2 (98.0 cM)였고, 중국밀과 미국밀 교배 집단에서는 보고된 4A 염색체에 위치하는
QSl.hwwgr-4AL (37.0 cM, 118.0 cM)도 본 연구에서 보고된 마커와 다른 위치였다(
Kumar et al. 2007,
Gao et al. 2015,
Guo et al. 2018,
Li et al. 2016,
2018). 앞에서 언급한 바와 같이 이 연구에서 사용된 표지인자는 SSR을 바탕으로 한 표지인자이다. 따라서, 향후 SNP나 sequencing을 통해 개발된 표지인자, 그리고 DNA chip과 같은 유전적 정보의 정확성이 높은 분석기법을 이용하여 국내 유전분석집단에 대한 추가적인 탐색이 이루어진다면 국내 육종 프로그램에서 수량 증진을 위한 긴 이삭 품종 선발의 효율성 증진이 가능할 것이다.
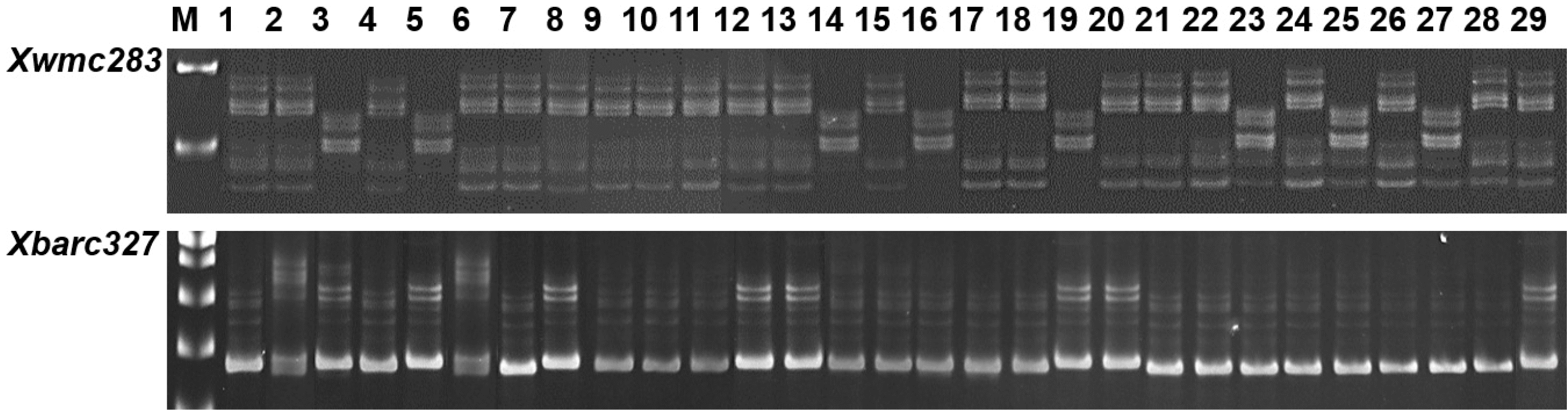
이 연구에서 확인된 QTL과 상응하는 표지인자를 이용하여 국내 밀 29품종에 대하여 표지인자의 유효성 검정 한 결과(
Table 3,
Fig. 4), 올그루와 유전적으로 상동성을 보인
a타입과 금강과 상동성을 보인
b타입, 그리고 전혀 다른 타입을 보인
c타입으로 구분되었다.
Xwmc283에 의해 국내 29 품종 중, 21 품종(72.4%)이
a타입을 보였으며, 8 품종(27.6%)이
b타입을 보였으며, 각각 이삭 길이는 7.68과 7.33 cm로 차이가 없었다.
Xbarc327에 의해 국내 8 품종(27.6%)이
a타입으로 19 품종(65.5%)이
b타입으로 확인되었으며, 평균 이삭 길이는 각각 7.60과 7.48 cm로 차이가 없었지만, 은파와 안백은 금강이나 올그루와 다른 유전자형을 나타내었고, 이들의 이삭 길이는 8.45 cm로 길었다. QTL의 유전자형을 이용하여 국내 29개 품종의 수장을 비교한 결과(
Table 4),
Xwmc283-
Xbarc327에 대한 유전자형에 따라 5개의 조합(
aa,
ab,
ac,
ba,
bb)으로 나타났고, 올그루와 같은 유전자형(
aa)을 지닌 품종들의 평균 이삭 길이는 7.86 cm로, 금강밀과 같은 유전자형(
bb)을 지닌 품종들은 7.42 cm보다 길었다. 올그루밀과 금강밀의 유전자형이 혼재된 품종들중에서
Xwmc283-Xbarc327의 유전자형이 올그루와 금강인 품종들(
ab)의 평균 이삭 길이가 7.51 cm로,
Xwmc283-Xbarc327의 유전자형이 금강과 올그루인 품종들(
ba)의 평균 이삭 길이가 7.17 cm로 보다 길었다.
Xwmc283의 유전자형은 올그루이면서
Xbarc327의 유전자형은 올그루와 금강과 다른(
ac) 은파와 안백은 8.45 cm로
ba와
bb형 보다 이삭 길이가 길었다. 반수체 집단에서 선발된
Xwmc283과
Xbarc327을 이용한 국내 품종에 대한 유효성 검정 한 결과,
Xbarc327의
c 형은 다른 타입보다 이삭 길이가 길었지만,
Xwmc283는 유전형 간에 차이가 없어서
Xbarc327가 보다 효과적인 표지인자로 활용이 가능할 것으로 보인다. 하지만 본 연구에 이용된 국내 반수체 밀 집단에 적용된 SSR 마커의 한계와 제한된 환경에서 수행된 특성 평가로 국내 육종프로그램에 직접적인 적용은 한계가 있기 때문에 기존에 보고된 이삭 길이 관련 표지인자(
Lee et al. 2014,
Kang et al. 2019)와 최근에 다양하게 수행되고 있는 SNP 검정을 통한 고밀도 유전지도 작성과 연계된 연구를 통하여 국내 품종의 이삭 길이 향상을 통한 수량 증대가 가능할 것이다.
적 요
국내 밀 품종의 수량성 향상을 위해 올그루와 금강을 모부본으로 만들어진 반수체 유전 집단에 대한 출수일수, 간장, 수장과 일수립수를 2년간 조사하였다. 총 1,482개의 SSR 마커를 검정하여 다형성을 보인 170개 SSR 마커를 이용하여 유전 및 QTL 분석을 실시하였다. 유전분석 결과, 출수일수, 간장 및 일수립수는 낮은 LOD 임계값을 나타내어 수장에 대해서만 유의성 있는 QTL 분석이 가능하였다. 염색체 4A에서 수장과 관련된 QTL이 측면표지인자 Xwmc283과 Xbarc327에 의하여 확인되었으며, 이 QTL의 표현형에 대한 기여도는 44.3%를 설명할 수 있는 주동유전자인 것으로 확인되었다. 국내 29개 밀 품종의 수장에 대한 유효성을 검정한 결과, Xwmc283은 국내 밀 품종을 2 그룹(a와 b)으로 분류하였으나 그룹간 수장의 차이는 보이지 않았다. Xbarc327은 국내 밀 품종을 3 그룹(a, b, 및 c)으로 분류하였으며 유전자형 ‘c’가 가장 긴 수장을 보였다. 두 측면표지인자의 조합은 최종적으로 국내 품종을 5 그룹(aa, ab, ac, ba, 및 bb)로 분류하였으며, 유전자형 ‘ac’가 가장 긴 수장을 보였다.
사 사
본 연구는 농촌진흥청 연구사업(세부과제명: 장수형 밀 농업 형질 개선을 위한 고밀도 유전자 지도 작성, 세부과제번호: PJ01279201)에 의해 이루어진 것임.
Fig. 1Frequency distributions of heading date (A), culm length (B), spike length (C) and kernel number per spike (D) in doubled haploid populations of Korean wheats. Arrows indicate mean value of parental cultivars.
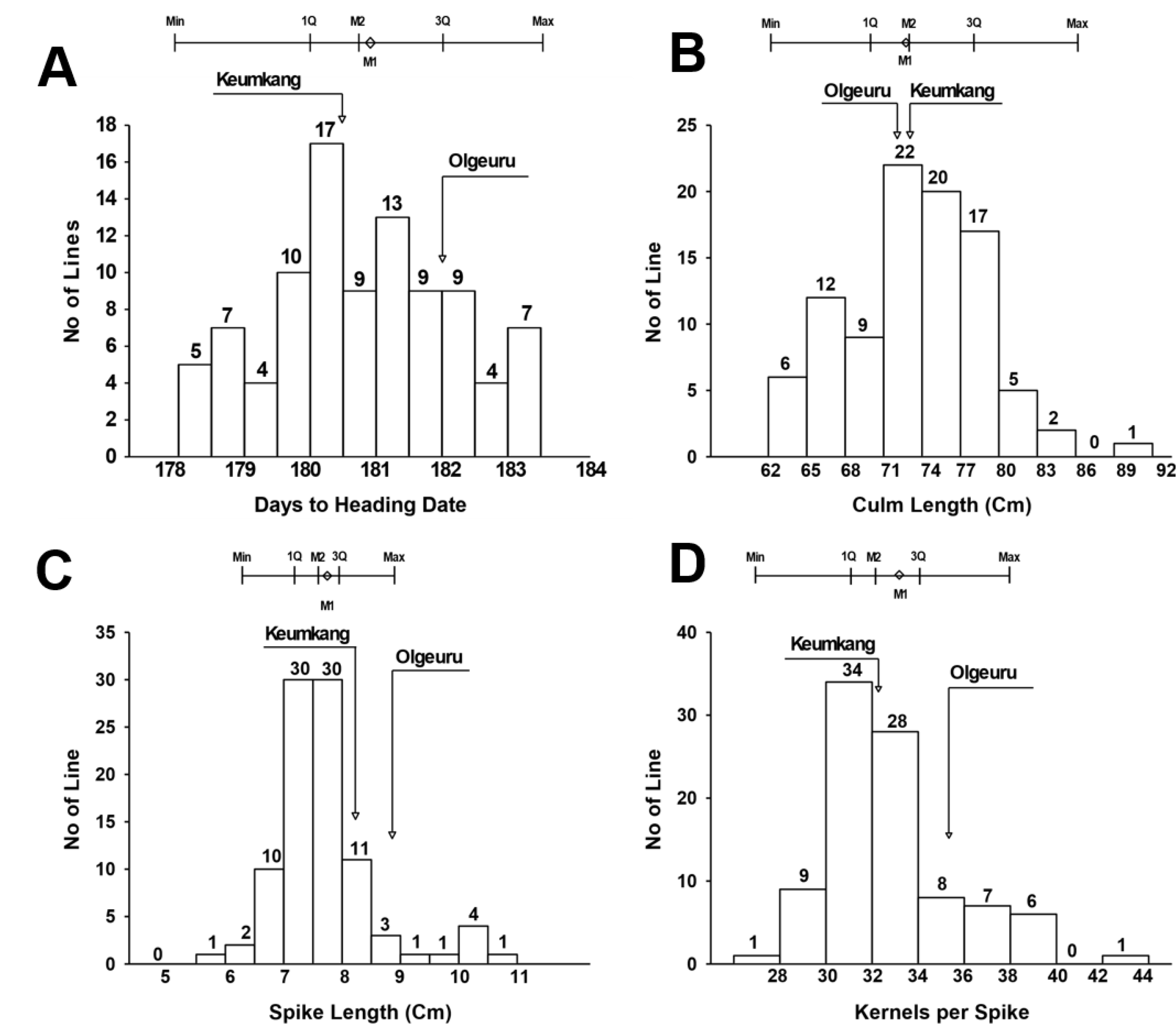
Fig. 2Expression of genotypic data by modified heat map of 94 doubled haploid populations of Korean wheats. Gray, black, bright gray color indicate Olgeuru (A type), Keumkang (B type), and hetero (C type), and white color indicates missing data. *, **, and *** significant at p≤0.05, p≤0.01, and p≤0.001 levels, respectively.

Fig. 3Linkage maps for spike length on chromosome 4A in of 94 doubled haploid populations of Korean wheats. Markers are indicated on the right and genetic distances (cM) are indicated on the left. QTLs for spike length (SL), qSL-1 was on chromosome 4A. LOD threshold (the dotted line), 3.37 was calculated by permutation (n=1,000) of IciMapping Ver. 4.0. HD=days to heading date, CL=culm length, SL=spike length, KNS=kernel number per spike.
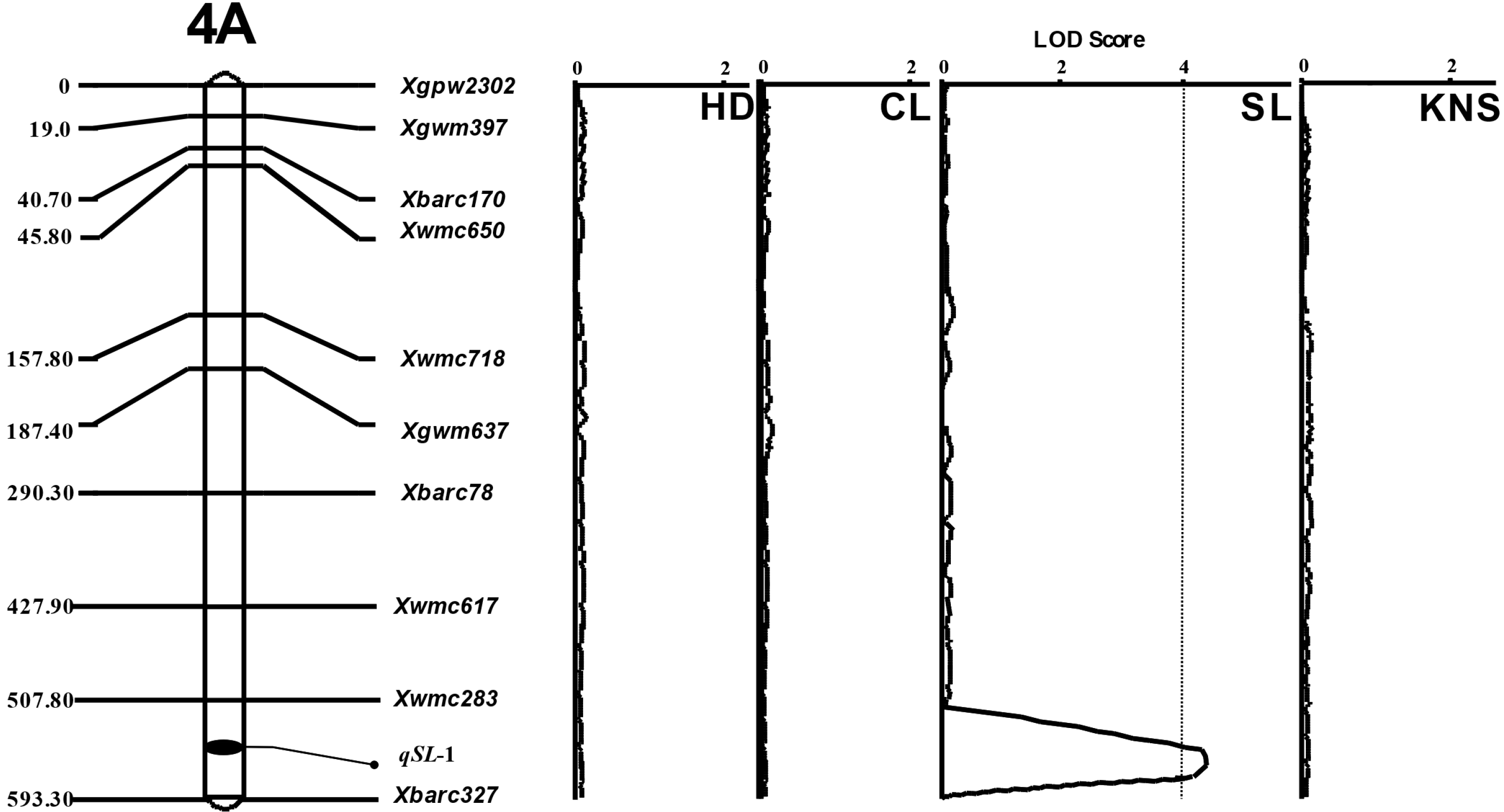
Fig. 4PCR products produced of QTL, Xwmc283 and Xbarc327, in 29 Korean wheat cultivars. 1, Ol; 2, Geuru; 3, Dahong; 4, Cheongkye; 5, Eunpa; 6, Tapdong; 7, Namhae; 8, Uri; 9, Olgeuru; 10, Alchan; 11, Gobun; 12, Keumkang; 13, Seodun; 14, Saeol; 15, Jipoom; 16, Milsung;17, Joeun; 18, Anbaek; 19, Jopoom; 20, Shinmichal; 21, Jonong; 22, Jokyung; 23, Younbaek; 24, Shinmichal1; 25, Baekjoong; 26, Jeokjoong; 27, Sukang; 28, Hanbaek; 29, Suan. a and b genotypes represent homozygous to Olgeuru and Keumkang, respectively. Genotype c means different type to a and b genotypes.
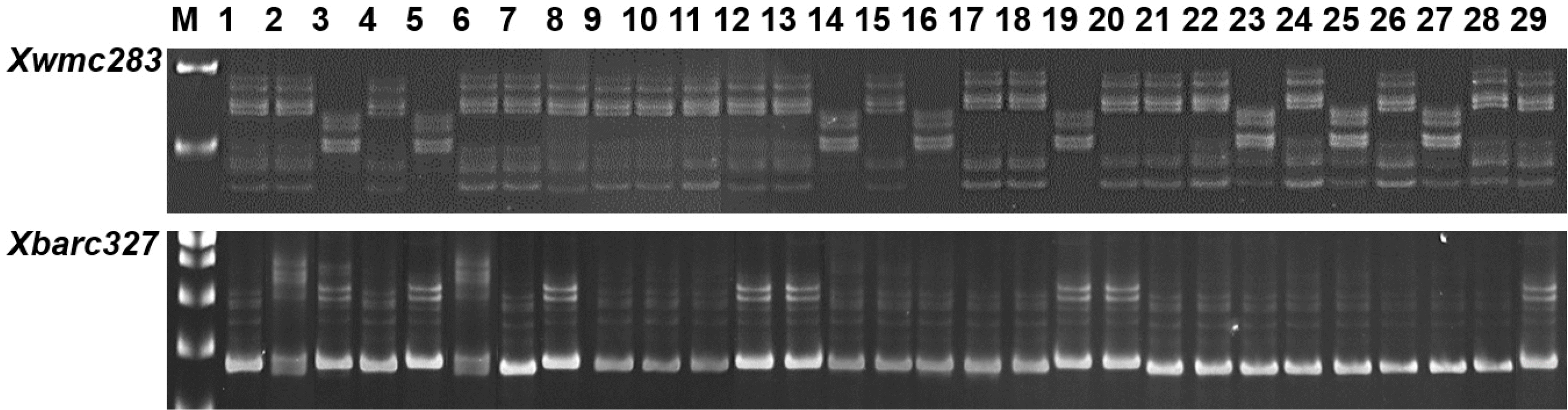
Table 1Descriptive statistics of 94 doubled haploid populations of Korean wheat.
Table 1
|
Agronomic traitsz
|
Descriptive statisticsy
|
|
|
Obs |
Mean |
Max |
Min |
Std |
Var |
Krt |
Skw |
|
HD |
94 |
180.9 |
183.5 |
178.0 |
1.4 |
1.9 |
-0.7 |
0.0 |
|
CL |
94 |
73.7 |
91.6 |
62.5 |
5.6 |
30.9 |
0.2 |
0.2 |
|
SL |
94 |
7.7 |
10.5 |
5.6 |
0.9 |
0.8 |
3.1 |
1.5 |
|
KNS |
94 |
32.9 |
42.5 |
27.1 |
2.9 |
8.2 |
0.7 |
0.8 |
Table 2QTLs for spike length using composite interval mapping in 94 doubled haploid populations of Korean wheat.
Table 2
|
QTL |
Chr. |
Left marker |
x2
|
Right marker |
x2
|
LODz
|
PVE (%)y
|
Additive effectx
|
|
qSL-1 |
4A |
Xwmc283
|
2.42ns
|
Xbarc327
|
90.04***w
|
4.06 |
44.3 |
1.16 |
Table 3Spike length according to the genotype of Xwmc283 and Xbarc327 in 29 Korean wheat cultivars for nine years under upland conditions.
Table 3
|
Cultivar |
Genotypesz
|
Spike Length (cm) |
|
|
Xwmc283
|
Xbarc327
|
|
Alchan |
a
|
b
|
6.8 |
|
Anbaek |
a
|
c
|
8.3 |
|
Baekjoong |
b
|
a
|
7.2 |
|
Cheongye |
a
|
b
|
7.4 |
|
Dahong |
b
|
a
|
6.8 |
|
Eunpa |
a
|
c
|
8.6 |
|
Geuru |
a
|
b
|
7.3 |
|
Gobun |
a
|
a
|
8.1 |
|
Hanbaek |
a
|
b
|
8.5 |
|
Jeokjoong |
a
|
b
|
7.0 |
|
Jinpoom |
a
|
b
|
8.5 |
|
Joeun |
a
|
a
|
6.8 |
|
Jokyung |
a
|
a
|
8.1 |
|
Jonong |
b
|
b
|
6.9 |
|
Jopoom |
a
|
b
|
6.7 |
|
Keumkang |
b
|
b
|
7.4 |
|
Milsung |
a
|
b
|
7.2 |
|
Namhae |
a
|
b
|
8.5 |
|
Ol |
b
|
a
|
7.5 |
|
Olgeuru |
a
|
a
|
9.1 |
|
Saeol |
a
|
b
|
7.1 |
|
Seodun |
a
|
b
|
7.8 |
|
Shinmichal |
b
|
b
|
7.9 |
|
Shinmichal1 |
a
|
b
|
7.9 |
|
Suan |
b
|
b
|
7.4 |
|
Sukang |
a
|
b
|
7.5 |
|
Tapdong |
b
|
b
|
7.5 |
|
Uri |
a
|
b
|
6.9 |
|
Younbaek |
a
|
a
|
7.2 |
Table 4Differences in spike length among Xwmc283 and Xbarc327 in 29 Korean wheat cultivars grown for nine years under upland conditions.
Table 4
|
Genotypea
|
No of cultivar |
Spike length (cm) |
|
Xwmc283
|
|
a
|
21 |
7.68ab
|
|
b
|
8 |
7.33a |
|
Xbarc327
|
|
a
|
8 |
7.60ab |
|
b
|
19 |
7.48b |
|
c
|
2 |
8.45a |
|
Xwmc283-Xbarc327
|
|
aa
|
5 |
7.86ab |
|
ab
|
14 |
7.51ab |
|
ac
|
2 |
8.45a |
|
ba
|
3 |
7.17b |
|
bb
|
5 |
7.42b |
References
- 1. Azadi A, Mardi M, Hervan EM, Abolghasem S, Mohammadi SA, Moradi F, Tabatabaee MT, Pirseyedi SM, Ebrahimi M, Fayaz F, Kazemi M, Ashkani S, Nakhoda B, Mohammadi-Nejad G. 2015. QTL mapping of yield and yield components under normal and salt-stress conditions in bread wheat (Triticum aestivum L.). Plant Mol Biol Rep 33: 102-120.
- 2. Bernardo R. 2008. Molecular markers and selection for complex traits in plants: Learning from the last 20 years. Crop Sci 48: 1649-1664.
- 3. Börner A, Schumann E, Fürste A, Cöster H, Leithold B, Röder MS, Weber WE. 2002. Mapping of quantitative trait loci determining agronomic important characters in hexaploid wheat (Triticum aestivum L.). Theor Appl Genet 105: 921-936.
- 4. Carter AH, Garland-Campbell K, Kidwell KK. 2011. Genetic Mapping of Quantitative Trait Loci Associated with Important Agronomic Traits in the Spring Wheat (Triticum aestivum L.) Cross 'Louise'×'Penawawa'. Crop Sci 51: 84-95.
- 5. Cho SW, Kang TG, Kang SW, Kang CS, Park CS. 2018. Assessment of DNA markers related to days to heading date, tiller number, and yield in Korean wheat populations. Korean J Breed Sci 50: 211-223.
- 6. Crossa J, Burgueño J, Dreisigacker S, Vargas M, Herrera-Foessel SA, Lillemo M, Singh RP, Trethowan R, Warburton M, Franco J, Reynolds M, Crouch JH, Ortiz R. 2007. Association analysis of historical bread wheat germplasm using additive genetic covariance of relatives and population structure. Genetics 177: 1889-1913.
- 7. Cui F, Ding A, Li J, Zhao C, Wang L, Wang X, Qi X, Li X, Li G, Gao J, Wang H. 2012. QTL detection of seven spike-related traits and their genetic correlations in wheat using two related RIL populations. Euphytica 186: 177-192.
- 8. Cuthbert JL, Somers DJ, Brûlé-Babel AL, Brown PD, Crow GH. 2008. Molecular mapping of quantitative trait loci for yield and yield components in spring wheat (Triticum aestivum L.). Theor Appl Genet 117: 595-608.
- 9. Deng S, Wu X, Wu Y, Zhou R, Wang H, Jia J, Liu S. 2011. Characterization and precise mapping of a QTL increasing spike number with pleiotropic effects in wheat. Theor Appl Genet 122: 281-289.
- 10. FAO. 2019. FAO crop production statistics. http://www.fao.org/faostat/en/#data/QC
- 11. Fischer AR. 2011. Wheat physiology: a review of recent developments. Crop Pasture Sci 62: 95-114.
- 12. Gao F, Wen W, Liu J, Rasheed A, Yin G, Xia X, Wu X, He Z. 2015. Genome-wide linkage mapping of QTL for yield components, plant height and yield-related physiological traits in the Chinese wheat cross Zhou 8425B/Chinese Spring. Front Plant Sci 6: 1099
- 13. Guo J, Shi W, Zhang Z, Cheng J, Sun D, Yu J, Li X, Guo P, Hao C. 2018. Association of yield-related traits in founder genotypes and derivatives of common wheat (Triticum aestivum L.). BMC Plant Biology 18: 38
- 14. Hai L, Guo H, Wagner C, Xiao S, Friedt W. 2008. Genomic regions for yield and yield parameters in Chinese winter wheat (Triticum aestivum L.) genotypes tested under varying environments correspond to QTL in widely different wheat materials. Plant Sci 175: 226-232.
- 15. Heidari B, Sayed-Tabatabaei EB, Saeidi G, Kearsey M, Suenaga K. 2011. Mapping QTL for grain yield, yield components, and spike features in a doubled haploid population of bread wheat. Genome 54: 517-27.
- 16. Hou J, Jiang Q, Hao C, Wang Y, Zhang H, Zhang X. 2014. Global selection on sucrose synthase haplotypes during a century of wheat breeding. Plant Physiol 164: 1918-1929.
- 17. Huang XQ, Cloutier S, Lycar L, Radovanovic N, Humphreys DG, Noll JS, Somers DJ, Brown PD. 2006. Molecular detection of QTLs for agronomic and quality traits in a doubled haploid population derived from two Canadian wheats (Triticum aestivum L.). Theor Appl Genet 113: 753-766.
- 18. Inagaki MN, Mujeeb-Kazi A. 1995. Comparison of polyhaploid production frequencies in crosses of hexaploid wheat with maize, pearl millet and sorghum. Korean J Breed Sci 45: 157-161.
- 19. Jantasuriyarat C, Vales MI, Watson CJW, Riera-Lizarazu O. 2004. Identification and mapping of genetic loci affecting the free-threshing habit and spike compactness in wheat (Triticum aestivum L.). Theor Appl Genet 108: 261-273.
- 20. Kang SW, Kim KM, Kang TG, Kang CS, Chun JB, Park CS, Cho SW. 2019. Mapping of QTL for yield traits in recombinant inbred lines derived from Korean wheat with long spike length. Korean J Breed Sci 52: 9-19.
- 21. Kumar N, Kulwal PL, Balyan HS, Gupta PK. 2007. QTL mapping for yield and yield contributing traits in two mapping populations of bread wheat. Mol Breed 19: 163-177.
- 22. Lee HS, Jung JU, Kang CS, Heo HY, Park CS. 2014. Mapping of QTL for yield and its related traits in a doubled haploid population of Korean wheat. Plant Biotechnol Rep 8: 443-454.
- 23. Li C, Bai G, Carver BF, Chao S, Wang Z. 2016. Mapping quantitative trait loci for plant adaptation and morphology traits in wheat using single nucleotide polymorphisms. Euphytica 208: 299-312.
- 24. Li F, Wen W, He Z, Liu J, Jin H, Cao S, Geng H, Yan J, Zhang P, Wan Y, Xi X. 2018. Genome-wide linkage mapping of yield-related traits in three Chinese bread wheat populations using high-density SNP markers. Theor Appl Genet 128: 1903-1924.
- 25. Li F, Wen W, Liu J, Zhang Y, Cao S, He Z, Rasheed A, Jin H, Zhang C, Yan J, Zhang P, Wan Y, Xi X. 2019. Genetic architecture of grain yield in bread wheat based on genome-wide association studies. BMC Plant Biol 19: 168
- 26. Li WL, Nelson JC, Chu CY, Shi LH, Huang SH, Liu DJ. 2002. Chromosomal locations and genetic relationships of tiller and spike characters in wheat. Euhytica 125: 357-366.
- 27. Liu J, Xu Z, Fan X, Zhou Q, Cao J, Wang F, Ji G, Yang L, Feng B, Wang T. 2018. A genome-wide association study of wheat spike related traits in China. Front Plant Sci 9: 1584
- 28. Liu L, Wang L, Yao J, Zheng Y, Zhao C. 2010. Association mapping of six agronomic traits on chromosome 4A of wheat (Triticum aestivum L.). Mol Plant Breed 1: 1-10.
- 29. Lozada DN, Mason RE, Babar MA, Carver BF, Guedira GB, Merrill K, Arguello MN, Acuna A, Vieira L, Holder A, Addison C, Moon DE, Miller RG, Dreisigacker S. 2017. Association mapping reveals loci associated with multiple traits that affect grain yield and adaptation in soft winter wheat. Euphytica 213: 222
- 30. Ma L, Li T, Hao C, Wang Y, Chen X, Zhang X. 2015. TaGS5-3A, a grain size gene selected during wheat improvement for larger kernel and yield. Plant Biotechnol J 14: 1269-1280.
- 31. Ma Z, Zhao D, Zhang C, Zhang Z, Xue S, Lin F, Kong Z, Tian D, Luo Q. 2007. Molecular genetic analysis of five spike-related traits in wheat using RIL and immortalized F2 populations. Mol Gen Genet 277: 31-42.
- 32. MAFRA2019. Ministry of Agriculture, Food and Rural Affairs statistical yearbook.
- 33. Marza F, Bai GH, Carver BF, Zhou WC. 2006. Quantitative trait loci for yield and related traits in the wheat population Ning7840×Clark. Clark. Theor Appl Genet 112: 688-698.
- 34. McCartney CA, Somers DJ, Humphreys DG, Lukow O, Ames N, Noll J, Cloutier S, McCallum BD. 2005. Mapping quantitative trait loci controlling agronomic traits in the spring wheat cross RL4452×'AC Domain'. AC Domain'. Genome 48: 870-883.
- 35. Nadolska-Orczyk A, Rajchel IK, Orczyk W, Gasparis S. 2017. Major genes determining yield-related traits in wheat and barley. Theor Appl Genet 130: 1081-1098.
- 36. Parry MAJ, Reynolds M, Salvucci ME, Raines C, Andralojc PJ, Zhu XG, Price GD, Condon AG, Furbank RT. 2011. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J Exp Bot 62: 453-467.
- 37. Quarrie SA, Quarrie SP, Radosevic R, Rancic D, Kaminska A, Barnes JD, Leverington M, Ceoloni C, Dodig D. 2006. Dissecting a wheat QTL for yield present in a range of environments: from the QTL to candidate genes. J Exp Bot 57: 2627-2637.
- 38. Quarrie SA, Steed A, Calestani C, Semikhodskii A, Lebreton C, Chinoy C, Steele N, Pljevljakusic' D, Waterman E, Weyen J, Schondelmaier J, Habash DZ, Farmer P, Saker L, Clarkson DT, Abugalieva A, Yessimbekova M, Turuspekov Y, Abugalieva S, Tuberosa R, Sanguineti MC, Hollington PA, Arague' R, Royo A, Dodig D. 2005. A high-density genetic map of hexaploid wheat (Triticum aestivum L.) from the cross Chinese Spring×SQ1 and its use to compare QTLs for grain yield across a range of environments. Theor Appl Genet 110: 865-880.
- 39. RDA2012. Standard of research and analysis for agricultural technology.
- 40. Reynolds MP, Foulkes MJ, Furbank R, Griffiths S, King J, Murchie M, Parry M, Slafer G. 2012. Achieving yield gains in wheat. Plant Cell Enviorn 35: 1799-1823.
- 41. Simmonds J, Scott P, Leverington-Waite M, Turner AS, Brinton J, Korzun V, Snape J, Uauy C. 2014. Identification and independent validation of a stable yield and thousand grain weight QTL on chromosome 6A of hexaploid wheat (Triticum aestivum L.). BMC Plant Biol 14: 191
- 42. Son JH, Kang CS, Cheong YK, Kim KH, Kim HS, Park JC, Kim KH, Kim BK, Park CS. 2015. Characterization of Korean wheat line with long spike. I. Agronomic traits and genetic variations. Korean J Breed Sci 47: 219-228.
- 43. Sukumaran S, Lopes M, Dreisigacker S, Reynolds M. 2018. Genetic analysis of multi-environmental spring wheat trials identifies genomic regions for locus-specific trade-offs for grain weight and grain number. Theor Appl Genet 131: 985-998.
- 44. Sun C, Zhang F, Yan X, Zhang X, Dong Z, Cui D, Chen F. 2017. Genome-wide association study for 13 agronomic traits reveals distribution of superior alleles in bread wheat from the Yellow and Huai Valley of China. Plant Biotech J 15: 953-969.
- 45. Valluru R, Reynolds MP, Salse J. 2014. Genetic and molecular bases of yield-associated traits: A translational biology approach between rice and wheat. Theor Appl Genet 127: 1463-1489.
- 46. Wang RX, Hai L, Zhang XY, You GX, Yan CS, Xiao SH. 2009. QTL mapping for grain filling rate and yield-related traits in RILs of the Chinese winter wheat population Heshangmai×Yu8679. Theor Appl Genet 118: 313-325.
- 47. Wang S, Yan X, Wang Y, Liu H, Cui D, Chen F. 2016. Haplotypes of the TaGS5-A1 gene are associated with thousand-kernel weight in Chinese bread wheat. Front Plant Sci 7: 783
- 48. Wang S, Zhang X, Chen F, Cui D. 2015. A single-nucleotide polymorphism of Tags5 gene revealed its association with kernel weight in Chinese bread wheat. Front Plant Sci 6: 1166
- 49. Ward BP, Brown-Guedira G, Kolb FL, Van Sanford DA, Tyagi P, Sneller CH, Griffey CA. 2019. Genome-wide association studies for yield related traits in soft red winter wheat grown in Virginia. PLoS ONE 14: e0208217.
- 50. Wu X, Cheng R, Xue S, Kong Z, Wan H, Li G, Huang Y, Jia H, Jia J, Zhang L, Ma Z. 2014. Precise mapping of a quantitative trait locus interval for spike length and grain weight in bread wheat (Triticum aestivum L.). Mol Breed 33: 129-138.
- 51. Würschum T, Leiser WL, Langer SM, Tucker MR, Longin CFH. 2018. Phenotypic and genetic analysis of spike and kernel characteristics in wheat reveals long-term genetic trends of grain yield components. Theor Appl Genet 131: 2071-2084.
- 52. Yao J, Wang L, Liu L, Zhao C, Zheng Y. 2009. Association mapping of agronomic traits on chromosome 2A of wheat. Genetica 137: 67-75.
- 53. You Q, Yang X, Peng Z, Xu L, Wang J. 2018. Development and applications of a high throughput genotyping tool for polyploid crops: single nucleotide polymorphism (SNP) array. Front Plant Sci 9: 104
- 54. Zhai H, Feng Z, Du X, Song Y, Liu X, Qi Z, Song L, Li J, Li L, Peng H, Hu Z, Yao Y, Xin M, Xiao S, Sun Q, Ni Z. 2018. A novel allele of TaGW2-A1 is located in a finely mapped QTL that increases grain weight but decreases grain number in wheat (Triticum aestivum L.). Theor Appl Genet 131: 539-553.
- 55. Zhang K, Tian J, Zhao L, Liu B, Chen G. 2009a. Detection of quantitative trait loci for heading date based on the doubled haploid progeny of two elite Chinese wheat cultivars. Genetica 135: 257-265.
- 56. Zhang K, Zhang Y, Chen G, Tian J. 2009b. Genetic analysis of grain yield and leaf chlorophyll content in common wheat. Cereal Res Comm 37: 499-511.
- 57. Zhang LY, Liu DC, Guo XL, Yang WL, Sun JZ, Wang DW, Zhang A. 2010a. Genomic distribution of quantitative trait loci for yield and yield-related traits in common wheat. J Integr Plant Biol 52: 996-1007.
- 58. Zhang ZB, Xu P, Jia JZ, Zhou RH. 2010b. Quantitative trait loci for leaf chlorophyll fluorescence traits in wheat. Aust J Crop Sci 4: 571-579.
- 59. Zhou Y, Conway B, Miller D, Marshall D, Cooper A, Murphy P, Chao S, Brown-Guedira G, Costa J. 2017. Quantitative trait loci mapping for spike characteristics in hexaploid wheat. Plant Genome 10: doi:10.3835/plantgenome2016.10.0101.