적요
Wheat (Triticum aestivum L.) is a major cereal crop grown worldwide, providing approximately 20% calorie and 25% protein intake. Wheat productivity is significantly affected by high temperatures, particularly during the grain-filling period. Heat stress accelerates leaf senescence, impairs photosynthesis, reduces starch accumulation, and alters protein synthesis, ultimately leading to a decrease in grain yield and quality. To mitigate the adverse effects of heat stress, wheat utilizes adaptation mechanisms, including the expression of heat shock proteins, activation of antioxidant defense systems, osmotic regulation, and transcription factor-mediated gene regulation. Stay-green traits also play a role in maintaining photosynthetic efficiency at high temperatures. Breeding strategies such as traditional breeding, marker-assisted selection , genomic selection , and genome editing are being explored to improve heat tolerance. Recent advances in the CRISPR-Cas9 technology enable precise gene editing, thereby enhancing the resilience of wheat to heat stress. Additionally, quantitative trait locus mapping and genome-wide association studies facilitated the identification of genetic regions associated with heat tolerance, thereby accelerating the development of climate-resilient wheat varieties. Future research should focus on integrating genetic and molecular approaches with sustainable agronomic practices and crop modeling strategies to optimize wheat productivity under rising temperatures. The integration of advanced breeding techniques and improved crop management can facilitate the development of wheat varieties that are more resilient to climate change.
서언
고온 스트레스는 밀의 생리⋅형태⋅생화학적 변화 과정에 광범위한 영향을 미친다(
Barnabás et al. 2008,
DuPont & Altenbach 2003,
Mirosavljević et al. 2021a). 주요 영향으로는 막 유동성 변화, 광합성 저하, 엽록소 함량 감소로 인한 잎의 노화, 단백질 변성 등이 있으며, 이는 생산량과 품질 저하로 이어진다(Asthir et al. 2015b,
Khan et al. 2020). 특히, 등숙기 고온은 등숙 기간 단축, 효소 활성 변화 등을 유발하여 전분과 단백질의 축적 감소로 인해 생산성을 저해한다(
Asseng et al. 2011,
Hafeez et al. 2023,
Mirosavljević et al. 2021a,
Zhao et al. 2007). 밀은 고온 스트레스에 반응하고 적응하기 위해 뿌리 발달 조절을 통해 수분과 영양분을 효율적으로 흡수하고(
Vincent & Gregory 1989,
Zhang et al. 2010), 기공 개폐 조절을 통해 증산 속도를 조절하여 수분 손실을 최소화한다(
Martínez-Goni et al. 2024,
Tricker et al. 2018). 고온 스트레스에 의한 세포 손상을 최소화하기 위해 항산화 방어 시스템을 활성화하여 ROS 축적을 억제하고(
Caverzan et al. 2016), 삼투조절 물질을 축적하여 세포 내 수분 균형을 유지한다(
Darko et al. 2019,
Qaseem et al. 2019). 또한, 열충격단백질(Heat Shock Proteins, HSPs) 발현을 통해 단백질 변성을 억제하고, 세포막의 유동성을 조절하여 막 손상을 방지한다(
Bourgine & Guihur 2021,
Haslbeck & Vierling 2015,
Horváth et al. 2008). 분자적 수준에서 DREB (Dehydration Responsive Element Binding proteins) (
Morran et al. 2011,
Niu et al. 2020), HSF (Heat Shock Factors) (
Guo et al. 2020,
Kumar et al. 2018,
Xue et al. 2015), NAC (NAM, ATAF1/2, and CUC2) (
Guérin et al. 2022,
Guo et al. 2015), WRKY (
Cheng et al. 2021,
El-Esawi et al. 2019) 등의 전사인자가 고온 스트레스에 반응하여 하위 유전자들을 발현 조절을 통해 고온에 대한 내성을 부여한다.
국내 밀 재배 환경에서 등숙기 중 기온 상승이 생산성과 품질에 영향을 미칠 것으로 예상되지만, 고온에 의한 국산 밀 품종의 생리적 반응 및 수량⋅품질에 대한 연구는 아직 부족한 실정이다. 따라서 고온 저항성 기작을 규명하기 위해 형태적, 생리적, 생화학적 반응을 종합적으로 고려한 연구가 필요하다. 본 논문에서는 밀의 성장과 발달에 대한 고온 스트레스 반응을 중심으로, 형태⋅생리⋅생화학적 변화, 고온 적응 매커니즘, 그리고 고온 스트레스 완화 방법을 논의하고자 한다. 이를 통해 기후 변화에 대응하기 위한 등숙기 고온 스트레스 연구 방향을 모색하고자 한다.
밀의 고온 스트레스 반응
형태학적 반응 및 생육 변화
생리적 반응 변화
잎의 노화
밀의 잎 노화는 sink 구조가 형성된 후 발달 중인 종실로 자원이 재이동하면서 발생하는 자연스러운 과정이나, 고온에서는 이 과정이 가속화되어 이용 가능한 자원의 양이 제한되며, 결국 수확량 감소로 이어진다(
Thomas & Howarth 2000,
Tuinstra et al. 1997). 고온(>34℃) 환경에서는 엽록소 생합성이 억제되어 잎의 노화가 촉진된다(
Akter & Rafiqul Islam 2017,
Paulsen 1994). 또한, 엽록소 분해가 가속화되고, 틸라코이드 막 손상 등의 요인으로 인해 엽록소 함량이 감소한다(
Ristic et al. 2007). 이러한 엽록소의 손실은 잎의 노화를 더욱 촉진하는 요인이 된다(
Liu & Huang 2000). 고온에서는 잎의 노화와 엽록소 손실이 빠르게 진행되어 종실로의 동화산물 공급이 감소하고 등숙이 저하되며 이로 인해 등숙 기간이 단축될 수 있다(
Shirdelmoghanloo et al. 2016). 등숙기 고온 환경에서는 잎의 노화가 촉진되고 종실의 호흡 속도가 빨라지면서 탄수화물의 이용 가능성이 감소한다. 이로 인해 종실 무게가 줄어들어 수확량에도 영향을 미친다(
Al-Khatib & Paulsen 1984,
Zhao et al. 2007).
Shirdelmoghanloo et al. (2016)의 연구에 따르면, 등숙 초기(10 days after anthesis, DAA)에 3일간 고온(37℃/27℃)을 처리했을 때, 지엽의 엽록소 감소가 종실 무게 감소와 가장 높은 상관관계를 나타냈으며, 이는 짧은 기간의 고온에서도 엽록소의 안정성이 종실 무게 유지에 중요한 요인이 될 수 있음을 시사한다. 또한,
Zhao et al. (2007)은 일교차가 큰 고온 환경에서 지엽의 노화가 더욱 촉진된다고 보고한 바 있다. 결국, 고온 환경에서는 엽록소 손실과 잎의 노화가 빠르게 진행되어 동화산물 공급이 감소하고, 이로 인해 종실 무게가 줄어들어 밀의 생산성 저하를 초래한다.
종실형태
Zhao et al. (2022)에 따르면, 고온(35℃/22℃) 환경에서는 립폭이 립장보다 더욱 크게 감소하며, 전반적으로 더 작은 종자가 형성되는 경향을 보였다.
Wang et al. (2018a)은 고온(37℃/17℃)에서 립장은 유의한 영향을 받지 않는 반면, 립폭이 감소한다고 보고하였다.
Rangan et al. (2019) 또한 유사한 결과를 보고하였는데, 등숙 초기(11~14 DAA)와 후기(27~30 DAA)에 3일간 고온(38℃/20℃)에 노출될 경우 천립중, 립폭, 종실 경도가 감소하지만, 립장에는 유의한 영향을 미치지 않는 것으로 나타났다. 이는 립장이 고온 처리 이전의 등숙 단계에서 이미 결정되었기 때문으로 판단된다(
Rangan et al. 2019). 또한, 같은 연구에서 내고온성 품종(Fang-60)은 천립중이 증가하는 경향을 보였으나, 립폭과 종실 경도에는 유의한 변화가 없었다. 반면, 고온에 민감한 품종에서는 A-type 전분 입자와 아밀로펙틴 집적 패턴이 변화하여 립폭과 종실중이 감소하였다(
Li et al. 2018).
Dias et al. (2008)은 고온(31℃/20℃) 환경에서 호분층과 배유세포의 구조 변화로 인해 종실이 수축되고, 그 결과 모양이 불량한 종자가 형성된다고 보고하였다. 이러한 연구 결과들은 고온 환경이 종실의 크기, 무게 및 구조적 특성에 영향을 미치며, 품종에 따라 내고온성 여부에 따른 차이가 존재함을 시사한다.
등숙 기간과 종실중
따라서, 밀의 생산성을 유지하기 위해서는 등숙기의 온도 변화를 면밀히 모니터링하고, 고온 환경에서도 등숙 속도를 효과적으로 증가시킬 수 있는 내고온성 품종을 개발하는 것이 중요하다.
생리⋅생화학적 반응 및 변화
광합성과 광계 시스템
광합성은 고온 스트레스에 가장 민감한 생리적 과정으로 광합성의 감소는 밀의 생육과 수확량에 직접적인 영향을 미친다(
Al-Khatib & Paulsen 1990,
1999,
Farooq et al. 2011). 고온 스트레스에 의한 광합성의 감소는 잎의 노화 촉진, 엽록소 함량 감소, 광합성 기구 손상(
Liu et al. 2017,
Mathur et al. 2014) 등을 유발하며, 이는 Ribulose-1,5-bisphosphate (RuBP) carboxylase/oxygenase (RubisCO)와 Rubisco activase (RCA)의 활성 감소가 원인이 된다(
Ristic et al. 2009). Rubisco는 50℃에서도 활성을 유지해 고온에서도 안정적이나(
Salvucci & Crafts-Brandner 2004a), RCA가 고온에서 불안정하여 Rubisco 활성이 저하된다(
Crafts-Brandner & Salvucci 2000,
Lal et al. 2022,
Salvucci & Crafts- Brandner 2004b).
Feng et al. (2014)의 연구에 따르면 겨울 밀 품종의 등숙기 고온은 Rubisco 활성과 엽록소 함량을 감소시키며, 엽록체와 미토콘드리아의 미세구조 손상을 유발하여 Rubisco 활성저해와 잎의 노화에 영향을 미친다. 또한 등숙 초기(10 DAA) 고온 (34℃/26℃, 16일)은 엽록소 함량과 광합성능을 감소시켜 수확량에 부정적인 영향을 미친다(
Pradhan & Prasad 2015).
고온에서 광합성 감소는 전자 전달 사슬의 핵심 구성요소인 광계Ⅱ (Photosystem Ⅱ, PSⅡ)와 관련이 있으며, 동계작물인 밀의 PSⅡ는 벼⋅수수 등 하계 작물보다 고온에 더욱 취약하다(
Al-Khatib & Paulsen 1999). 고온은 ROS의 생성을 유도하여 PSⅡ의 D1 단백질을 분해하고 비활성화 시키며(
Dwivedi et al. 2018,
Yamamoto et al. 2008), 동시에 틸라코이드 막의 유동성을 증가시켜 PSⅡ와 광수용 복합체Ⅱ (Light Harvesting Complexes Ⅱ, LHCⅡ) 간의 결합을 약화시킨다(
Gounaris et al. 1984,
Mathur et al. 2014). PSⅡ는 30~42℃의 온도에서 전자 전달이 감소하고, 산소 발생 복합체(Oxygen-Evolving Complex, OEC)와 관련된 효소의 비활성화로 인해 RuBP 재생 속도가 느려진다(
Salvucci & Crafts- Brandner 2004c). 등숙기에 고온(32℃/22℃)이 지속되면 엽록소 지수와 광합성 속도가 각각 25% 감소하며, 틸라코이드 막 손상은 68% 증가하고 기공전도도는 42% 늘어 수확량이 정상 조건 대비 44% 줄어드는 것으로 나타났다(
Djanaguiraman et al. 2020). 엽록소 형광 매개변수인 PSⅡ 최대 양자수율(Fv/Fm)은 내고온성 유전자형 스크리닝에 활용할 수 있는 지표로, 등숙 중기의 고온(38℃/28℃)에서 엽록소 지수와 Fv/Fm이 각각 평균적으로 44%, 63% 감소하였다(
Mirosavljević et al. 2024). 같은 연구에서 고온에서 엽록소 지수와 Fv/Fm이 종실중 및 수량과 정의 상관을 보였으며, 내고온성 품종은 고온에 민감한 품종에 비해 높은 엽록소 지수 및 Fv/Fm을 보였다(
Mirosavljević et al. 2024).
호흡작용
산화 스트레스
고온 환경에서는 밀의 생육에 영향을 미치는 ROS가 축적된다. 이러한 ROS에는 singlet oxygen (
1O
2), superoxide (O
2-), hydrogen peroxide (H
2O
2), hydroxyl radicals (⋅HO) 등이 포함되며, 세포 내에서는 ROS 생성과 소거가 균형을 이루어 산화환원 항상성을 유지한다(
Caverzan et al. 2016,
Poudel & Poudel 2020). 그러나 고온 환경에서 ROS가 항산화 능력을 초과하여 축적될 경우 산화스트레스가 발생하며(
Mullineaux & Baker 2010), 이는 세포 구조와 기능에 심각한 손상을 초래할 수 있다. 특히, 고온은 막 손상을 유발하여 전해질 누출을 증가시키고(
Savicka & Škute 2010), 미토콘드리아와 엽록체의 전자전달 효율을 감소시켜 ROS 축적을 유발한다(
Asthir et al. 2015a,
Mittler et al. 2022). 이로 인해 대사과정의 불균형이 초래되며(
Awasthi et al. 2015,
Barnabás et al. 2008,
Mittler 2002), 세포막 과산화가 증가하고 막의 열 안정성이 감소한다(
Bavita et al. 2012,
Savicka & Škute 2010). 또한, 세포막 탈분극, 지질 과산화, 단백질 산화, 효소 기능 저하, 핵산의 산화적 손상이 발생하여 세포예정사 경로가 활성화된다(
Poudel & Poudel 2020,
Sharma et al. 2012). ROS는 주로 엽록체, 미토콘드리아, 퍼옥시좀과 같은 산화적 대사가 활발한 세포소기관에서 생성되며(
Mittler et al. 2004), 특히 엽록체의 PSⅠ과 PSⅡ 반응 중심이 주요 ROS 생성 부위로 알려져 있다(
Asada 2006,
Soliman et al. 2011). 또한 고온 스트레스는 respiratory burst oxidase homologues (RBOHs)와 같은 ROS 생성 관련 효소를 암호화하는 유전자의 발현을 증가시켜 ROS 축적을 가속화한다(
Caverzan et al. 20l6).
고온 스트레스를 받은 밀은 세포막 지질 과산화로 인해 malondialdehyde (MDA) 농도가 증가하며, proline 및 betaglycine과 같은 삼투조절 물질이 축적되는 등 다양한 생리적 변화를 보인다(
Djanaguiraman et al. 2018,
Gupta et al. 2013,
Kumar et al. 2019,
Lal et al. 2022,
Savicka & Škute 2010). 밀의 유묘 단계에서 온도 상승에 따라 proline 농도는 감소하는 반면, H
2O
2 농도는 증가하는 경향을 보이며(
Kumar et al. 2012), 극단적인 고온(42℃)에서는 O
2- 생성 증가로 인해 MDA 농도가 급격히 상승한다(
Savicka & Škute 2010).
Chunduri et al. (2021)의 연구에 따르면, 개화 후 2일(2 DAA)부터 고온(35℃/17℃)을 처리한 밀은 7 DAA에서 대조구(24℃/17℃) 대비 proline과 H
2O
2 함량이 각각 238%, 29% 증가하였으며, 주야간 고온 조건(35℃/24℃)에서는 각각 780%, 61%까지 증가하였다. H
2O
2 농도 증가는 ROS 축적을 유도하여 신호전달 연쇄반응을 활성화하며(
Lamaoui et al. 2018), 이에 따라 Guaiacol peroxidase (GPOX) 활성화와 proline 함량 증가와 같은 방어 메커니즘이 작동하여 세포 내 H₂O₂ 농도를 조절함으로써 산화 스트레스에 대응하는 것으로 보고되었다(
Chunduri et al. 2021).
생산성
등숙기의 고온이 종실중에 미치는 영향은 처리 시기, 지속 기간, 품종의 유전적 특성에 따라 다르게 나타난다(
Hafeez et al. 2023). 특히, 등숙 초기의 고온 스트레스는 배유 세포 형성에 영향을 미쳐 종실중 감소를 유발하며, 등숙 후기의 고온은 광동화산물의 부족과 전분 생합성 효소의 비활성화를 초래하여 전분 합성을 저해한다(
Nicolas et al. 1985,
Sharma et al. 2019,
Yang et al. 2004).
Tashiro & Wardlaw (1989)의 연구에 따르면, 등숙 초기(7 DAA)부터 성숙기까지 고온(36℃/31℃)에 노출될 경우, 대조구 대비 종실 무게가 최대 85%까지 감소하였다. 또한, 14 DAA에 고온(32℃/22℃, 2주) 처리 시 종실중과 수량이 각각 39%와 44% 감소한다고 보고하였다(
Djanaguiraman et al. 2020).
Mirosavljević et al. (2021a,
2021b)의 연구에서도 14 DAA에 7일간 고온(38℃/28℃)에 노출될 경우, 수량과 종실중이 유의하게 감소하는 경향을 보였고,
Zhao et al. (2022)의 연구에 따르면 15~17 DAA 동안 고온(35℃/22℃)에서 수량 감소폭이 가장 큰 반면, 31~33 DAA의 고온 처리에서는 상대적으로 감소폭이 적었다.
이러한 연구 결과들은 등숙기의 고온이 종실중과 수확량 감소에 미치는 영향을 보여주며, 특히 고온의 발생 시기와 지속 시간이 중요한 요인임을 시사한다.
품질 변화
전분 생합성 및 함량
전분과 단백질의 축적은 등숙 초기에 결정되는 배유 세포의 수, 등숙 기간 및 속도에 영향을 받는 배유 세포의 최종 크기에 의해 결정된다(
Ullah et al. 2022). 전분은 종실 무게의 65~75%를 차지하며, 수량과 가공식품의 식미에 중요한 영향을 미친다(
Baik 2010,
Hurkman et al. 2003). 그러나 등숙기의 고온은 전분 합성을 저해하여 밀가루 생산량을 감소시키고, 아밀로스/아밀로펙틴 비율 변화, 전분 입자 크기 분포 변화 등을 초래하여 전분의 기능적 특성을 변화시킨다(
Hurkman et al. 2003,
Sharma et al. 2019). 배유에서 전분이 차지하는 비중이 높기 때문에, 고온에 의한 전분 축적 감소는 수량 감소로 이어진다(
Barnabás et al. 2008,
Jenner 1991,
Wang et al. 2018). 등숙기 고온에 따른 전분, 아밀로스, 아밀로펙틴의 함량 변화는
Table 2에 정리되어 있다.
밀에서 전분 합성은 포도당-1-인산을 전구체로 하여 다양한 대사 효소의 조절을 통해 이루어진다. 아밀로스는 ADP-glucose pyrophosphorylase (AGPase)와 granule-bound starch synthase (GBSS)에 의해 합성되며, 아밀로펙틴은 AGPase, soluble starch synthases (SSS), starch branching enzymes (SBE), starch debranching enzymes (DBE)에 의해 합성된다(
Jeon et al. 2010). 전분 생합성 유전자의 발현은 고온의 영향을 받으며, 단백질 합성보다 더욱 민감하게 반응하는 것으로 알려져 있다(
Bhullar & Jenner 1985,
Dupont & Altenbach 2003). 특히, SSS의 최적 활성 온도는 20~25℃로, 고온에서 발현이 크게 감소하고 활성이 저해된다(
Hawker & Jenner 1993,
Hurkman et al. 2003,
Prakash et al. 2004). 25℃ 이상의 고온에서는 SSS의 활성 저하로 인해 종실 발달이 저해되며(
Jenner et al. 1991,
Keeling et al. 1993,
Prakash et al. 2004), 30~40℃에서는 배유의 전분 축적이 약 30% 감소한다(
Jenner 1994,
Stone & Nicolas 1995,
Ullah et al. 2022). 특히, 40℃에서는 SSS의 활성이 97%까지 감소하여 기능을 거의 상실하게 된다(
Jenner 1994). 국산 밀 품종인 백강과 조경을 이용한 국내 연구에서도 고온 환경에서 전분 합성 관련 유전자 발현은 대조구에 비해 빠르게 감소하였다(
Cho et al. 2022). 고온 환경에서 전분 함량 감소는 주로 아밀로펙틴 감소로 인해 발생하며, 아밀로스 함량은 상대적으로 영향을 적게 받아 아밀로펙틴:아밀로스 비율이 낮아진다(
Li et al. 2018,
Liu et al. 2011,
Yan et al. 2008,
Zhao et al. 2006).
Liu et al. (2023)에 따르면 등숙기 고온(35℃/25℃)에서 아밀로펙틴의 짧은 사슬 함량은 증가하고 아밀로스 함량은 감소하며, 아밀로펙틴의 구조와 전분 품질은 등숙 중기 고온에서 가장 큰 영향을 받았다.
밀의 전분 입자는 크기에 따라 A type (10~35 μm)과 B type (1~10 μm)으로 구분된다(
Chiotelli & Le Meste 2002). 전분 입자의 비율과 형태 변화는 반죽의 점도 및 가공 제품의 식미에도 영향을 미친다(
Park et al. 2009,
Peterson & Fulcher 2001). 전분 입자의 크기, 모양, 구조는 25℃ 이상의 고온 환경에서 변화하며(
Liu et al. 2011), 이에 따라 A type 입자의 비율도 영향을 받는다. 일부 연구에서는 고온이 A type 입자의 비율을 증가시킨다고 보고한 반면(
Blumenthal et al. 1995,
Hurkman & Wood 2011), 다른 연구에서는 감소하는 것으로 보고되었다(
Li et al. 2018,
Liu et al. 2011,
Lu et al. 2014,
Wang et al. 2012). 이러한 차이는 고온 처리 방법과 처리 시기에 따라 달라지는 것으로 판단된다(
Lu et al. 2014). 따라서 등숙기 고온은 전분 생합성 효소의 활성을 저하시켜 종실 발달을 방해하며, 이로 인해 수량과 품질에 부정적인 영향을 미친다.
단백질 생합성 및 함량
밀가루의 품질은 단백질의 양적 및 질적 특성에 의해 결정되며, 이는 가공적성과 반죽 특성에 직접적인 영향을 미친다. 밀의 주요 단백질인 글루텐은 단량체인 글리아딘과 중합체인 글루테닌으로 구성되며, 밀가루 반죽에 신장성과 탄력성을 부여하는 역할을 한다(
Wieser et al. 2023).
반죽 강도 저하는 단백질 함량 증가보다 글루테닌/글리아딘 비율 변화와 관련이 깊다(
Daniel & Triboi 2000,
DuPont et al. 2006,
Wardlaw et al. 2002). 고온에서는 글루테닌 합성이 감소하는 반면, 글리아딘 합성은 유지되거나 증가하는데(
Majoul et al. 2004,
Stone & Nicolas 1996), 이는 글리아딘 유전자가 5' heat shock elements를 포함하고 있어 고온(30~35℃) 환경에서도 발현이 증가하거나 유지되기 때문이다(
Blumenthal et al. 1990). 결과적으로, 글루테닌/글리아딘 비율이 감소하면서 반죽 강도가 약화된다(
Ciaff et al. 1996). 또한, 글루텐 구조 형성에 중요한 역할을 하는 이황화 이성질화 효소(disulfide isomerase)의 활성이 감소하면 고분자 글루테닌 중합체의 합성이 줄어들어 반죽 특성에 부정적인 영향을 미친다(
Blumenthal et al. 1991,
1995). 그러나 일부 연구에서는 고온이 단백질 조성과 밀가루 가공적성에 영향을 미치지 않는다고 보고하고 있으며, 품종에 따라 단백질 특성이 결정된다는 상반된 결과도 존재한다(
Castro et al. 2007,
Son et al. 2019).
결론적으로, 등숙기의 고온 환경은 밀 단백질 조성과 글루테닌/글리아딘 비율 변화로 인해 밀가루의 가공 적성과 반죽 강도에 영향을 미친다. 따라서 고온에서도 안정적인 품질을 유지할 수 있는 품종 개발이 필요하다.
밀의 고온 스트레스 적응 메커니즘
Heat Shock Proteins (HSPs) 방어 기작
고온 스트레스는 단백질의 합성과 접힘(folding) 과정에 영향을 미쳐 단백질 오접힘(misfolding) 및 응집(aggregation)을 유발하며, 이는 세포 기능에 심각한 영향을 미칠 수 있다(Yadav & Bhatt et al. 2024). HSPs는 분자샤페론으로 작용하여 단백질의 접힘, 조립, 운반 및 분해를 담당하며, 고온과 같은 환경 스트레스에서 세포막과 단백질을 안정화한다(
Wang et al. 2004). 또한, 변성과 응집을 방지하고 오접힘된 단백질의 재접힘(refolding)을 도와 고온 스트레스 반응을 완화하는 역할을 한다.
HSPs는 분자량과 서열 유사도에 따라 HSP100/ClpB, HSP90, HSP70, HSP60, HSP40 및 small HSP (sHSP)/HSP20 6개의 family로 분류되며(
Kumar et al. 2020), 서로 다른 기능을 수행한다(
Table 3). 밀에서는 총 753개의
HSP 유전자가 확인되었으며(
Kumar et al. 2020), 품종별 고온 반응 차이에 따라 발현 수준이 다르게 나타난다. 개화기부터 수확기까지 고온(34℃/10℃)을 처리한 결과, HSP81, HSP80, HSP70과 HSP20을 포함한 sHSP의 발현이 증가하였다(
Majoul et al. 2003, 2004). 특히, 15~17 DAA에서 급격한 고온(40℃/25℃)을 처리한 경우 15~30 kDa 범위의 sHSP 발현이 증가했으며, 내고온성 품종에서는 더욱 강하게 발현되었다(
Skylas et al. 2002). 또한,
Hurkman et al. (2009) 연구에서도 10 DAA에 고온(37℃/28℃) 처리 시 HSP16.9와 HSP70이 증가하였다. 이는 16.9 kDa HSP가 밀의 내고온성과 밀접한 관련이 있으며, 내고온성 품종 육종에서 유망한 마커로 활용될 가능성이 있음을 시사한다.
HSPs의 발현 조절은 heat shock transcription factors (HSFs)에 의해 조절된다. HSPs를 암호화하는 heat shock genes (HSGs)의 프로모터에는 heat shock element (HSE)가 존재하며, 이는 palindromic 서열로 구성된다(
Scharf et al. 2012). 정상 조건에서 HSFs는 세포질에서 단량체 상태로 HSP70과 결합하여 비활성화되어 있지만, 고온 환경에서는 HSP70과 분리되어 삼중체를 형성하고 핵으로 이동하여 HSE에 결합함으로써 HSP 발현을 촉진한다(
Usman et al. 2014,
Haider et al. 2022). 특히,
TaHSFA6f의 과발현은
TaHSP16.8,
TaHSP17,
TaHSP17.3,
TaHSP90.1-A1 등의 다양한
HSPs와
glutathione-S-transferase 등의 표적 유전자와 상호작용하여 밀의 내고온성을 향상시킬 수 있음이 밝혀졌다(
Xue et al. 2015). 이러한 연구 결과는 HSFs의 조절 메커니즘이 밀의 고온 스트레스 적응에서 중요한 역할을 한다는 것을 시사한다.
항산화 방어 기작
고온 환경에서는 ROS의 생성이 증가하며, 이에 대응하여 식물의 항산화 방어 시스템이 활성화된다(
Kumar et al. 2012). 항산화 방어 시스템은 ROS를 제거하여 세포의 산화환원 균형을 유지하고(
Caverzan et al. 2016), 산화적 손상을 최소화하는 역할을 한다(
Poudel & Poudel 2020,
Xin et al. 2019). 항산화 방어 시스템은 효소적 항산화 작용과 비효소적 항산화 작용으로 구분된다(
Mittler 2002,
2004). 효소적 항산화 작용에는 superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), peroxidase (POD), glutathione peroxidase (GPX), glutathione reductase (GR), glutathione
S-transferase (GST), monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR) 등의 효소가 관여하며, 비효소적 항산화 작용에는 ascorbate (AsA), glutathione (GSH), tocopherol, carotenoids 등이 포함된다(
Caverzan et al. 2016,
Noctor & Foyer 1998). 고온 환경에서 생성된 O₂⁻은 SOD에 의해 O₂와 H₂O₂로 전환되며, H₂O₂는 CAT와 APX 등에 의해 분해된다(
Gill & Tuteja 2010,
Kaushal et al. 2016). 특히 CAT는 H₂O₂를 빠르게 H₂O와 O₂로 분해하며, APX는 Halliwell-Asada 경로(AsA-GSH cycle)를 통해 H₂O₂를 제거한다(
Asthir et al. 2015b,
Sofo et al. 2015). GPX도 APX와 유사한 역할을 수행하지만, GSH를 직접 환원제로 사용하여 H₂O₂를 제거한다(
Apel & Hirt 2004). 항산화 작용 매커니즘과 밀에서 고온에 의한 항산화 효소의 활성 변화는
Table 4에 정리되어 있다.
밀에서 SOD와 CAT의 활성은 고온 스트레스 초기에는 증가하지만, 등숙이 진행되면서 점차 감소하며, 이는 종실중 감소와 관련이 있다(
Jing et al. 2020,
Zhao et al. 2007). 일반적으로 항산화 효소의 활성은 특정 온도까지 증가하다가 이후 감소하는 경향을 보인다(
Hasanuzzaman et al. 2013). 내고온성 품종은 고온에 민감한 품종보다 항산화 효소 활성이 높아 산화적 손상이 적으며(
Almeselmani et al. 2006,
2009,
Asthir 2015a,
Goyal & Asthir 2010), 더 높은 온도에서도 효소 활성을 유지할 수 있다(
Chakraborty & Pradhan 2011). 등숙기 고온(37℃)에 노출된 내고온성 품종은 민감성 품종보다 광합성 효율의 감소가 적고, MDA 함량이 낮으며, CAT와 APX 활성이 높았다(
Zmani et al. 2018). 또한, 고온 처리 후에도 비교적 오랫동안 항산화 효소 활성 수준을 유지하는 특징을 보였다(
Zmani et al. 2018). 내고온성 품종은 민감성 품종보다 세포막 손상이 적고 APX 활성이 높으며, 광합성과 항산화 관련 단백질의 발현이 증가하여 산화적 손상을 완화한다(
Wang et al. 2015). 이러한 연구 결과들은 밀의 고온 스트레스 적응에 항산화 방어 시스템이 중요한 역할을 하며, 특히 내고온성 품종이 높은 항산화 효소 활성을 통해 산화적 스트레스를 완화하고 생산성 감소를 방지하는 주요 기작임을 시사한다. 이는 내고온성 품종 개발과 환경적 관리 전략 수립에 중요한 기초 자료가 될 수 있다.
Stay-green (SG)
Stay-green (SG)은 개화 후에도 잎의 노화가 지연되어 스트레스 조건에서도 엽록소 함량과 광합성 능력을 유지하는 특성으로, 식물의 생산성과 환경 적응성 향상에 중요한 역할을 한다(
Rosenow 1983,
Thomas & Howarth 2000). SG 특성은 크게 기능적 SG와 비기능적 SG로 구분되며, 기능적 SG는 노화의 시작을 지연시키는 유형(type A)과 노화 속도를 더디게 하여 광합성 능력을 유지하는 유형(type B)으로 나뉜다. 반면, 비기능적 SG는 광합성 능력이 감소하나 엽록소 분해 경로가 손상되어 엽색이 유지되는 유형(type C)으로 분류된다(
Thomas & Howarth 2000). SG는 잎의 광합성을 지속하고 등숙 기간을 연장함으로써 수확량을 증가시키며, 특히 고온 스트레스 환경에서 작물의 생육 안정성을 높이는 중요한 형질이다(
Fokar et al. 1998,
Kumari et al. 2013,
Reynolds et al. 2000). 연구 결과에 따르면, SG 특성을 가진 밀 유전자형은 일반 품종보다 수량과 강한 양적 상관을 보였으며(
Kumari et al. 2013), 특히 고온 환경에서도 엽록소 함량 유지와 등숙률 및 종실중 증가 등의 특징을 나타냈다(
Zarei et al. 2013). 이 외에도 많은 연구에서 SG 특성이 고온 환경에서 작물 수확량을 증가시키는 중요한 요인으로 작용한다는 결과들이 확인되었다(
Latif et al. 2020,
Pinto et al. 2016,
Roy et al. 2021,
Vignjevic et al. 2015).
SG 특성은 canopy 온도 감소와도 밀접한 연관이 있다.
Kumari et al. (2013)에 따르면, SG 유전자형에서 canopy temperature depression (CTD) 값이 유의미하게 높게 나타났으며, 이는 잎이 오래 유지되면서 지속적인 증산작용을 통해 canopy 온도가 낮아지는 냉각 효과 때문으로 해석된다(
Fischer et al. 1998,
Reynolds et al. 1994). 이러한 특성은 식물의 생리적 활력을 유지하여 극한 환경에서도 광합성 효율을 높이는 데 중요한 역할을 한다. 따라서 SG 특성은 내고온성 품종 육성에 중요한 형질로 간주되며, 이를 활용한 작물 개량을 위해 SG 관련 유전자 및 QTL 탐색 등의 연구가 필요하다.
전 세계적인 기후 변화로 인해 농업 생산성이 지속적으로 위협받고 있으며, 특히 고온 스트레스는 밀 생산과 식량 안보에 심각한 영향을 미치고 있다. 이를 해결하기 위해 내고온성 품종 육종, 수자원 및 토양 관리, 작물 관리 기술 등의 전략이 필수적이다. 특히, 식물의 형태학적, 생리⋅생화학적, 분자적 반응을 연구하여 고온 스트레스에 대한 적응 메커니즘을 이해하고, 이를 기반으로 교배 육종, 분자육종, 유전체 선발, 유전자 교정 등 다양한 방법을 활용한 내고온성 품종 개발이 이루어지고 있다(
Janni et al. 2020,
Kim et al. 2018,
Zhang et al. 2019).
전통 육종
또한, 재래종 밀(Landraces)은 유전적 변이가 풍부한 6배체(Hexaploid) 밀과 교배하기 용이하고(
Trethowan & Mujeeb- Kazi 2008), 극한 환경에서도 적응할 수 있는 유전적 형질을 보유하고 있다(
Kumar et al. 2023). 예를 들어, 스페인의 재래종 밀에서 선발된 ‘Aragon 03’은 고온 내성이 강해 현재까지 재배되고 있으며(
Royo & Briceño-Félix 2011), 야생 엠머밀(
T. turgidum)은 듀럼밀(
T. durum)과 빵밀(
T. aestivum)의 A/B 게놈을 공유하여 고온 스트레스 내성과 관련된 유전적 특성을 제공할 수 있다(
Peng et al. 2013).
Molero et al. (2023)의 연구에서는 재래종 및 근연야생종을 활용한 봄밀 자원 149개 중, 일부 유래 계통이 고온에서도 높은 수확량을 보였다. 또한,
Gudi et al. (2024)의 연구에서는 빵밀(
T. aestivum)의 D 게놈 공여체인
Aegilops tauschii 유래 도입 계통(introgression lines, ILs) 369점 중 고온과 가뭄에 내성을 지닌 계통을 선발하여, 내고온성 관련 유전적 배경을 육종 소재로 활용할 가능성을 제시하였다. 그러나 이러한 육종 방법은 시간과 노력이 많이 들기 때문에 내고온성 관련 유전적 배경을 효율적으로 전달하기 위해서 유전적 기작 연구가 수반되어야 한다(
Gudi et al. 2024). 빵밀 품종에서는 ‘Stay-green’ 형질이 고온 내성과 밀접한 관련이 있어(
Nawaz et al. 2013), 내고온성 품종 육종을 위한 육종 재료로 활용할 수 있을 것으로 판단된다.
Lu et al. (2022)의 연구에서는 유묘기 내고온성을 가진 계통이 성체에서도 내고온성을 유지하는 경향을 보여, 유묘기 단계에서 조기 선발을 통해 육종 기간을 단축할 수 있는 가능성을 제시하였다.
분자표지 기반(Marker-Assisted Selection, MAS) 육종
마커 보조 선발(MAS)은 원하는 형질을 가진 식물을 효율적으로 선발하여 작물의 생산성, 병 저항성 및 스트레스 내성을 향상시키는 기술이다(
Ashraf et al. 2012). MAS 기반 기술에는 마커 보조 역교배(Marker-Assisted Backcrossing, MABC), 마커 보조 반복 선발(Marker-Assisted Recurrent Selection, MARS), 전장 유전체 연관분석(Genome-Wide Association Studies, GWAS) 등이 포함되며, 이러한 방법들은 전통적인 육종 방식보다 더 빠르고 정밀하게 유용한 형질을 선발할 수 있도록 한다(
Singha et al. 2022). 특히, GWAS와 QTL (Quantitative Trait Locus) 분석은 고온 스트레스 내성과 관련된 유전적 특성을 밝히는 데 중요한 역할을 하며, 이를 통해 내고온성 품종 개발을 위한 연구가 활발히 진행되고 있다(
Paliwal et al. 2012).
또한, QTL 매핑을 통해 밀의 고온 내성 기작을 규명하고, 이를 기반으로 고온 환경에서도 우수한 생육을 보이는 품종 개발이 이루어지고 있다(
Langridge & Reynolds 2015,
Paliwal et al. 2012).
Bhusal et al. (2017)에 따르면 밀의 고온 스트레스 내성과 관련된 QTL이 수량과 수량구성요소 관련 형질에서 확인되었으며, 특히 일수립수, 천립중, 등숙률과 등숙 기간과 밀접한 연관성이 있는 것으로 확인되었다. 이러한 연구 결과는 내고온성 밀 품종 개발을 위한 유전적 지표로 활용될 수 있다. 따라서 MAS 기반 유전체 육종 기술은 내고온성 밀 품종 개발을 가속화하는 효과적인 방법이며, 이를 통해 기온 상승으로 인한 밀 생산성 감소를 최소화하고, 안정적인 곡물 공급을 가능하게 할 것으로 판단된다.
유전체 선발(Genomic Selection, GS) 육종
유전체 선발(Genomic Selection, GS)은 DNA 마커를 활용하여 다양한 형질에 대한 개체의 육종 가치를 예측하는 육종 기법으로, 새로운 품종을 개발하는데 효과적이다(
Tayade et al. 2018). 특히, 밀 유전체의 복잡성과 다양한 생물적⋅비생물적 스트레스에 대한 내성이 강화된 품종 개발의 필요성이 높아지면서 GS가 주목받고 있다. 고밀도 SNP array의 개발로 인해 GS의 적용이 더욱 용이 해졌으며, MAS와 비교하여 작은 효과를 가지는 대립유전자의 탐지가 가능하고 마커 선택 과정에서 활용이 가능하다는 장점이 있다(
Cairns & Prasanna 2018). 또한, GS는 육종 집단에서 복합 형질에 대한 육종 가치를 예측하여 유용한 유전 자원을 선발하는데 활용될 수 있으며, 복잡한 형질의 유전적 개량을 강화하는데 기여한다(
Bhat et al. 2016,
Wang et al. 2018)
유전체 교정(Genome Editing) 육종
밀은 6배체의 복잡한 유전체를 가진 작물로 고온 내성을 비롯한 다양한 형질에는 많은 유전자가 관여하며, 이러한 유전자들의 기능을 연구하기 위해 CRISPR-Cas9, TALENs, ZFNs 등의 유전체 교정 기술이 활용되고 있다(
Kim et al. 2018). 이 중 CRISPR-Cas9 시스템은 수량 증가, 품질 개선, 제초제 저항성, 생물적 및 비생물적 스트레스 내성 향상 등 다양한 분야에서 널리 활용되고 있다(
Hussain et al. 2018) (
Table 5).
밀에서는 CRISPR-Cas9을 활용하여 생산성 향상, 품질 개선, 저장성 증대 및 글루텐 저감과 같은 다양한 연구들이 수행되었다. 생산성 향상을 위해 밀의 종실 형태와 종실중에 영향을 미치는
TONNUAU 1과 이삭 배열에 관련된 유전자를 교정하여 수량을 증가시키는 연구가 수행되었다(
Wang et al. 2019,
Wolde et al. 2019). 또한, 질소 이용 효율을 높이고 품질과 생육 특성을 개선하기 위한 유전자 교정 연구가 수행되었으며(
Gupta et al. 2023,
Zhang et al. 2021a,
Zhang et al. 2021b),
TaARF15-A1 변이를 통해 밀의 노화를 억제하는 연구가 수행되었다(
Li et al. 2023). 이와 함께, ω- 및 γ-글리아딘 유전자 클러스터를 교정함으로써 글루텐 함량을 감소시키고 면역반응성을 줄이는 연구도 수행되었다(
Jouanin et al. 2020,
Yu et al. 2024). 저장성과 식품 안전성을 개선하기 위해 아스파라긴 축적을 감소시키는 연구가 진행되었으며(
Raffan et al. 2021),
Tamyb10을 교정하여 수발아 저항성이 높은 적립게 밀을 개발하는 연구도 수행되었다(
Zhu et al. 2023).
한편, 밀의 고온 내성을 강화하기 위한 연구도 활발히 이루어지고 있다. 고온 내성을 조절하는 주요 표적 유전자로
HSTF (
Heat Shock Transcription Factor)와
RCA (
Rubisco Activase)가 연구되고 있다. 고온 환경에서
HSTF의 교정을 통해 HSPs의 발현을 증가시켜 밀의 고온 내성을 강화할 수 있으며, 광합성 과정에서 Rubisco 활성을 유지하는
RCA 교정을 통해 광합성 효율과 내고온성을 개선할 수 있다(
Debbarma et al. 2019). 이와 함께, 밀의 수분 이용 효율을 개선하기 위해 기공 개폐를 조절하는 유전자를 교정하여 고온 환경에서도 효율적인 수분 사용이 가능하도록 조절할 수 있다. 유전자 편집 기술의 발전을 통해 밀의 생산성 향상, 품질 개선, 저장성 증대 및 내고온성 품종 개발이 더욱 정밀하고 효율적으로 이루어질 것으로 기대된다.
결론 및 향후 전망
밀은 전 세계적으로 중요한 작물이며, 기후 변화로 인해 등숙기 고온 스트레스가 생산성과 품질에 미치는 영향이 더욱 커지고 있다. 본 논문에서는 밀의 고온 스트레스 반응과 적응 메커니즘, 그리고 내고온성 품종 육종 전략에 대해 종합적으로 분석하였다. 고온 스트레스는 밀의 형태적, 생리적, 생화학적 변화를 유발하며, 주요 영향으로는 엽록소 함량 감소, 광합성 효율 저하, 전분 및 단백질 축적 감소 등이 있다. 등숙기 고온은 잎 노화를 가속화하고 등숙 기간을 단축시켜 수량과 품질 저하를 초래한다. 이에 대한 밀의 대응 기작으로 열충격단백질(HSPs) 발현 증가, 항산화 방어 시스템 활성화, 삼투 조절 물질 축적 등이 작용한다. 특히, Stay-Green (SG) 특성을 가진 품종은 광합성 능력을 유지하여 고온 스트레스에서도 생산성을 유지할 가능성이 높다.
내고온성 밀 품종 개발을 위해 전통 육종, 분자 마커 기반 육종(MAS), 유전체 선발(GS), 유전체 교정(GE) 등 다양한 전략이 활용되고 있다. 특히, CRISPR-Cas9과 같은 유전자 편집 기술은 내고온성 유전자의 발현을 조절하여 고온 스트레스 내성을 향상시키는 강력한 도구로 평가된다. 또한, 고온 스트레스 반응 관련 유전자 및 QTL 탐색을 통해 내고온성 품종 개발이 가속화될 것으로 기대된다.
향후 연구에서는 고온에 대한 작물의 반응을 더욱 정밀하게 이해하기 위한 유전자 수준의 연구와 환경 스트레스 내성을 고려한 지속 가능한 농업 시스템 구축이 필요하다. 또한, 기후 변화에 대응하는 작물 모델링 연구 및 최적의 재배 관리 기술 개발이 병행되어야 할 것이다. 이를 통해, 밀의 생산성을 유지하고, 고온 스트레스에 대한 대응 전략을 마련하여 글로벌 식량 안보를 강화할 수 있을 것으로 전망된다.
적요
밀(Triticum aestivum L.)은 전 세계적으로 재배되는 주요 곡물로, 인류에게 약 20%의 열량과 25%의 단백질을 공급한다. 그러나 밀의 생산성은 특히 등숙기 동안의 고온으로 인해 크게 영향을 받는다. 고온 스트레스는 잎의 노화를 촉진하고, 광합성을 저해하며, 전분 축적을 감소시키고, 단백질 합성을 변화시켜 결국 수량과 품질을 저하시킨다. 이를 극복하기 위해 밀은 열충격단백질(Heat Shock Proteins, HSPs) 발현, 항산화 방어 시스템 활성화, 삼투 조절, 전사인자 매개 유전자 조절 등의 적응 기작을 활용한다. 또한, Stay-Green (SG) 형질은 고온 환경에서도 광합성 효율을 유지하는 데 중요한 역할을 한다. 밀의 내열성을 향상시키기 위해 전통 육종, 마커 보조 선택(Marker-Assisted Selection, MAS), 유전체 선택(Genomic Selection, GS), 유전체 편집(Genome Editing, GE) 등의 육종 전략이 연구되고 있다. 특히, CRISPR-Cas9 기술의 발전은 표적 유전자 편집을 통해 밀의 내열성을 정밀하게 향상시키는 데 기여하고 있다. 아울러, 양적형질좌(QTL) 분석 및 전장 유전체 연관분석(GWAS)을 활용하여 고온 내성과 관련된 유전 영역을 규명함으로써 기후 변화에 강한 밀 품종 개발을 가속화할 수 있다. 향후 연구는 유전학적⋅분자적 접근법과 지속 가능한 농업 관리 전략, 작물 모델링 기법을 융합하여 기온 상승 속에서도 최적의 밀 생산성을 유지하는 데 초점을 맞춰야 한다. 고급 육종 기술과 개선된 작물 관리 전략을 통합하면 기후 변화에 적응할 수 있는 밀 품종 개발이 더욱 촉진될 것이다.
사사
본 논문은 농촌진흥청 연구사업(과제번호: PJ01723101)의 지원에 의해 이루어진 것임.
본 연구는 2025년도 농촌진흥청 국립식량과학원 전문연구원 과정 지원사업에 의해 이루어진 것임.
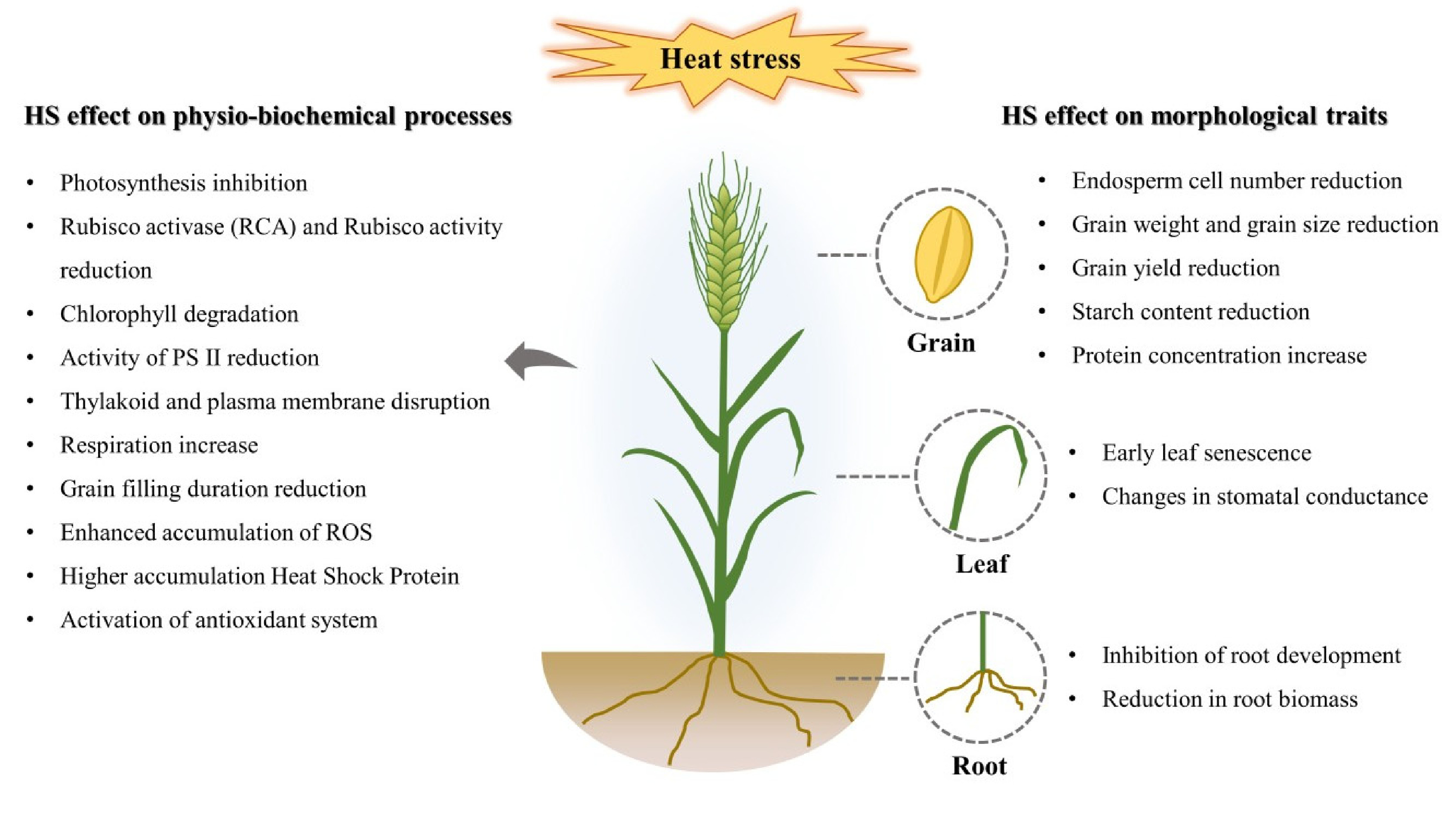
Fig. 1Schematic representation showing the heat stress effects on physio-biochemical processes and/or morphological traits in wheat.
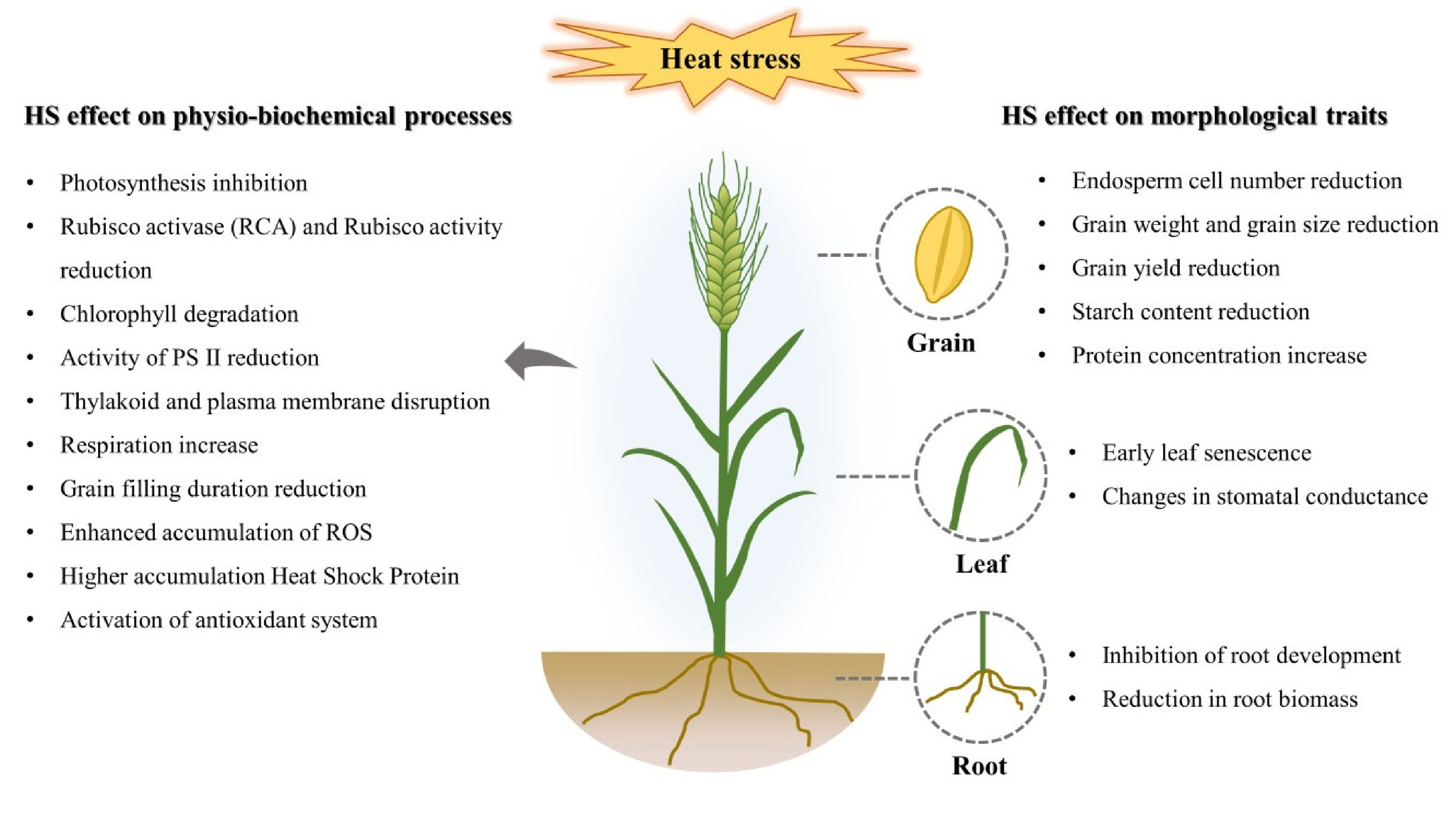
Table 1Effect of heat stress on grain filling duration (GFD), grain weight (GW), 1000-grain weight (TGW) and grain yield (GY) in wheat.
Table 1
|
Temperature |
Duration |
Cultivar |
GFD (day) |
GW (%) |
TGW (%) |
GY (%) |
Ref |
|
30℃, 35℃ or 40℃ |
6-38 DAAz(3 days interval) |
Yangmai 9 |
13↓ |
- |
- |
- |
Liu et al. 2011
|
|
Yangmai 12 |
15↓ |
- |
- |
- |
|
35℃/20℃ |
10-24 DAA |
Fleisch481 |
- |
19.1↓ |
17.1↓ |
- |
Bányai et al. 2014
|
|
Soissons |
- |
38.0↓ |
30.4↓ |
- |
|
Plainsman |
- |
35.9↓ |
34.2↓ |
- |
|
Magma |
- |
27.9↓ |
20.0↓ |
- |
|
20-34 DAA |
Fleisch481 |
- |
1.8↓ |
7.0↓ |
- |
|
Soissons |
- |
36.7↓ |
28.1↓ |
- |
|
Plainsman |
- |
23.8↓ |
26.9↓ |
- |
|
Magma |
- |
3.6↓ |
7.1↓ |
- |
|
35℃/26℃ |
14-19 DAA |
15 spring wheat |
3-12↓ |
2.6-41.9↓ |
- |
8.3-50.9↓ |
Vignjevic et al. 2015
|
|
37℃/17℃ |
15-18 DAA |
Chinese Spring |
8↓ |
20↓ |
- |
- |
Wang et al. 2018
|
|
38℃/20℃ |
11-14 DAA |
Banks |
- |
- |
16.2↓ |
- |
Rangan et al. 2019
|
|
EGA Gregory |
- |
- |
51.7↓ |
- |
|
Fang-60 |
- |
- |
16.8↑ |
- |
|
27-30 DAA |
Banks |
- |
- |
24.9↓ |
- |
|
EGA Gregory |
- |
- |
16.2↓ |
- |
|
Fang-60 |
- |
- |
NS |
- |
|
32℃/22℃ |
14-28 DAA |
Seri82 |
- |
39.0↓ |
- |
44.0↓ |
Djanaguiraman et al. 2020
|
|
35℃/25℃ & 38℃/28℃ |
0-7 DAA & 14-21 DAA |
12 winter wheat |
- |
38.5-68.5↓ |
23.0-51.4↓ |
- |
Mirosavljević et al. 2021a
|
|
38℃/28℃ |
14-21 DAA |
12 winter wheat |
- |
19.2-49.3↓ |
0.7-41.1↓ |
- |
|
35℃/22℃ |
7-33 DAA(3 days interval) |
Ningmai 13 |
2-7↓ |
- |
6.3-46.8↓ |
19.8-48.4↓ |
Zhao et al. 2022
|
|
Zhenmai 12 |
2-7↓ |
- |
3.5-47.7↓ |
13.9-52.0↓ |
|
38℃/28℃ |
mid-grain filling |
9 wheat varieties |
|
11.3-38.2↓ |
|
9.3-73.0↓ |
Mirosavljević et al. 2024
|
Table 2Effect of heat stress on starch and protein contents in wheat grain.
Table 2
|
Temperature |
Duration |
Cultivar |
Starch content (%) |
Amylose content (%) |
Amylopectin content
(%) |
Protein content (%) |
Gliadin content (%) |
Glutenin content (%) |
Ref |
|
39.8℃ |
5-9 DAAz
|
Jimai 20 |
4.0↓ |
0.9↑ |
5.0↓ |
- |
- |
- |
Yan et al. 2008
|
|
Lumai 21 |
1.5↓ |
0.9↑ |
2.5↓ |
- |
- |
- |
|
34℃/22℃ |
7-11 DAA |
Xuzhou 26 |
7.4↓ |
- |
- |
1.4↑ |
- |
- |
Zhao et al. 2008
|
|
Yangmai 9 |
14.5↓ |
- |
- |
0.8↑ |
- |
- |
|
32℃/24℃ |
7-11 DAA |
Xuzhou 26 |
3.6↓ |
- |
- |
1.8↑ |
- |
- |
|
Yangmai 9 |
9.0↓ |
- |
- |
1.6↑ |
- |
- |
|
34℃/30℃ |
7-14 DAA |
Yang9 |
9.2↓ |
2.3↓ |
7.0↓ |
- |
- |
- |
Wang et al. 2012
|
|
35℃/20℃ |
12-27 DAHy
|
Mv Magma |
- |
- |
- |
3.5↑ |
- |
- |
Rakszegi et al. 2014
|
|
Fatima 2 |
- |
- |
- |
3.8↑ |
- |
- |
|
Plainsman V |
- |
- |
- |
3.8↑ |
- |
- |
|
40℃/20℃ |
10-17 DAA |
Antequera |
- |
- |
- |
2.9↓ |
- |
- |
Tomás et al. 2020
|
|
Pata Negra |
- |
- |
- |
1.0↓ |
- |
- |
|
35℃/22℃ |
7-33 DAA(3 days interval) |
Ningmai 13 |
2.8-8.0↓ |
1.4↓
(7-9 DAA) |
2.1-7.4↓ |
0.3-3.9↑ |
0.3-0.9↑ |
0.2-0.7↑ |
Zhao et al. 2022
|
|
Zhenmai 12 |
1.8-4.5↓ |
0.4↓
(7-9 DAA) |
1.6-4.4↓ |
0.3-1.7↑ |
0.2-0.9↑ |
0.2-0.8↑ |
Table 3The function of heat shock proteins (HSPs) under heat stress in plants.
Table 3
|
HSPs category |
Major functions |
Reference |
|
sHSP |
HSP10 |
Assists in protein folding within mitochondria and chloroplasts, preventing misfolding under stress conditions. |
Kumar et al. 2020
|
|
HSP20 |
Prevent protein aggregation, enhance thermotolerance, and protect cellular components. |
Pandey et al. 2015
|
|
HSP40 |
Acts as a co-chaperone, regulating HSP70 activity, facilitating protein refolding, and preventing aggregation. |
Kumar et al. 2020
|
|
HSP60 |
Mitochondrial & chloroplast chaperonin; assists in protein folding and prevents misfolding under stress. |
Kumar et al. 2020
|
|
HSP70 |
Major chaperone; protects proteins from heat stress, assists in refolding and protein degradation. |
Heidari et al. 2024
|
|
HSP90 |
Protein folding, stabilizing stress-related proteins, and maintaining cellular homeostasis. |
Singh et al. 2024
|
|
HSP100 |
Disaggregates and refolds damaged proteins after severe heat stress; aids in recovery. |
Kumar et al. 2020
|
|
HSP101 |
Prevents protein aggregation, repairs denatured proteins, and plays a crucial role in heat stress recovery and tolerance. |
Erdayani et al. 2020
|
Table 4ROS scavenging reactions by different antioxidants (
Yadav et al. 2022) and changes of antioxidant activity under heat stress in wheat.
Table 4
|
Mechanism |
Antioxidants |
Major catalyzed reactions |
Temperature |
Duration |
Ref |
|
Enzymatic antioxidants |
Super oxide dismutase (SOD) |
2O2- + 2H+ → H2O2 + O2
|
35℃/25℃ |
45 days after germination-maturity |
Almeselmani et al. 2009
|
|
34℃/22℃ or 32℃/24℃ |
7-11 DAAz
|
Zhao et al. 2007
|
|
Field (late sown) |
- |
Kumar et al. 2023
|
|
Catalase (CAT) |
H2O2 → H2O + 1/2O2
|
35℃/20℃ |
12-27 DAHy
|
Balla et al. 2009
|
|
30℃, 35℃ or 40℃ |
6h (1-month-old seedling) |
Sarkar et al. 2016
|
|
field |
- |
Dwivedi et al. 2024
|
|
Glutathione peroxidase (GPX) |
2GSH + ROOH (H2O2) → GSSG + ROH + H2O (2H2O) |
35±1℃ |
48h (8-day-old seedling) |
Mohi-Ud-Din et al. 2021
|
|
Ascorbate peroxidase (APX) |
H2O2 + 2AsA → 2H2O + 2MDHA |
37℃/25℃ |
7 DAA-maturity |
Zmani et al. 2018
|
|
34℃/24℃ |
48h (six-leaf stage) |
Khanzada et al. 2024
|
|
Monodehydro ascorbate reductase (MDHAR) |
NADPH + H+ + 2MDHA → 2AsA + NADP+
|
Field (late sown) |
- |
Khanna-Chopra & Chauhan 2015
|
|
Dehydroascorbate reductase (DHAR) |
DHA + 2GSH → AsA + GSSG |
33±1℃ |
6 days (germinated seed) |
Goyal & Asthir 2010
|
|
Glutathione reductase (GR) |
NADPH + H+ + GSSG → 2GSH + NADP+
|
35℃/20℃ |
12-27DAH |
Balla et al. 2007
|
|
42℃ |
2h (7-day-old seedling) |
Gupta et al. 2013
|
|
Glutathione-S-transferase (GST) |
H2O2 + 2GSH → 2H2O + GSSG RX + GSH→ HX + GS-R |
35℃/20℃ |
12-27 DAH |
Balla et al. 2009
|
|
- |
- |
Wang et al. 2015
|
|
35±1℃ |
48h (8-day-old seedling) |
Mohi-Ud-Din et al. 2021
|
|
Non-enzymatic antioxidants |
Ascorbate (AsA) |
Scavenges O2-, H2O2, OH⋅, and O2
|
30℃ or 35℃ |
7 days (4-day-old seedling) |
Kumar et al. 2013
|
|
Glutathione (GSH) |
Scavenges H2O2, OH⋅, and O2
|
37℃ |
3 days (2 weeks seedling) |
Kocsy et al. 2002
|
|
Glutathione (GSH) |
Scavenges H2O2, OH⋅, and O2
|
35±1℃ |
48 h (8-day-old seedling) |
Mohi-Ud-Din et al. 2021
|
|
Tocopherol |
Scavenges O2, OH⋅, ROO⋅ and ROOH |
30℃ or 35℃ |
7 days (4-day-old seedling) |
Kumar et al. 2013
|
Table 5Genome editing for abiotic stress and agronomic traits improvement in wheat.
Table 5
|
Target gene |
Modification type |
Delivery technique |
Desired trait |
Reference |
|
TaMLO
|
Insertion |
Agrobacterium-mediatedtransformation |
Powdery mildew resistance |
Wang et al. 2014
|
|
TaDEP1, TaGW2
|
Base editing |
Biolistics based transformation |
Herbicide resistance |
Mishra et al. 2020
|
|
TaDREB2, TaERF3
|
Deletion or insertion, SNP |
Transient expression |
Drought stress |
Kim et al. 2018
|
|
TaFT-D1
|
Deletion |
Biolistics based transformation |
Increased number of spikelets |
Chen et al. 2022
|
|
TaGASR7
|
Deletion or insertion |
Transient expression |
NS |
Lin et al. 2020
|
|
TaGW2
|
Gene knockout |
Biolistics based transformation |
Enhanced seed size and grain weight |
Wang et al. 2018
|
|
TaCKX2-1, TaGLW7, TaGW2, TaGW8 |
Deletion or insertion |
Biolistics based transformation |
Grain regulation |
Zhang et al. 2019
|
|
TaASN2
|
Deletion |
Biolistics based transformation |
Reduced free asparagine |
Raffan et al. 2021
|
|
TaMYB10
|
Deletion |
Agrobacterium-mediated transformation |
Pre-harvest sprouting |
Zhu et al. 2023
|
|
TaSPL13
|
Deletion |
Agrobacterium-mediated transformation |
Flowering time, Tiller number |
Gupta et al. 2023
|
|
TaARF15
|
SNP |
Biolistics based transformation |
Leaf senescence |
Li et al. 2023
|
|
ω- and γ-gliadin gene clusters |
Deletion |
Biolistics based transformation |
Grain protein |
Yu et al. 2024
|
References
- 1. Abasi F, Raja NI, Mashwani ZR, Ehsan M, Ali H, Shahbaz M. 2024. Heat and wheat: Adaptation strategies with respect to heat shock proteins and antioxidant potential; an era of climate change. Int J Biol Macromol 256: 128379.
- 2. Akter N, Rafiqul Islam M. 2017. Heat stress effects and management in wheat. A review. Agron Sustain Dev 37: 1-17.
- 3. Al-Khatib K, Paulsen GM. 1984. Mode of high temperature injury to wheat during grain development. Physiol plantarum 61: 363-368.
- 4. Al-Khatib K, Paulsen GM. 1990. Photosynthesis and productivity during high-temperature stress of wheat genotypes from major world regions. Crop Sci 30: 1127-1132.
- 5. Al-Khatib K, Paulsen GM. 1999. High-temperature effects on photosynthetic processes in temperate and tropical cereals. Crop Sci 39: 119-125.
- 6. Allan RP, Arias PA, Berger S, Canadell JG, Cassou C, Chen D, Cherchi A, Connors SL, Coppola E, Cruz FA. 2023. Intergovernmental panel on climate change (IPCC). Summary for policymakers. In: Climate change 2021: The physical science basis. Contribution of working group I to the sixth assessment report of the intergovernmental panel on climate change. Cambridge University Press, pp. 3-32.
- 7. Almeselmani M, Deshmukh P, Sairam R. 2009. High temperature stress tolerance in wheat genotypes: Role of antioxidant defence enzymes. Acta Agron Hung 57: 1-14.
- 8. Almeselmani M, Deshmukh PS, Sairam RK, Kushwaha SR, Singh TP. 2006. Protective role of antioxidant enzymes under high temperature stress. Plant Sci 171: 382-388.
- 9. Apel K, Hirt H. 2004. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55: 373-399.
- 10. Asada K. 2006. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol 141: 391-396.
- 11. Ashraf M, Akram NA, Foolad MR. In: Shabala S, Cuin T. 2012. (Eds). Marker-assisted selection in plant breeding for salinity tolerance. . Plant salt tolerance: Methods in molecular biology. Humana Press, Totowa, NJ. pp. 305-333,
- 12. Asseng S, Foster IAN, Turner NC. 2011. The impact of temperature variability on wheat yields. Global Change Biol 17: 997-1012.
- 13. Asthir B. 2015a. Protective mechanisms of heat tolerance in crop plants. J Plant Interact 10: 202-210.
- 14. Asthir B. 2015b. Mechanisms of heat tolerance in crop plants. Biol Plantarum 59: 620-628.
- 15. Awasthi R, Bhandari K, Nayyar H. 2015. Temperature stress and redox homeostasis in agricultural crops. Front Environ Sci 3: 11
- 16. Baik BK. Hou GG (Ed.).2010. Effects of flour protein and starch on noodle quality. . Asian noodles science, technology, and processing. John Wiley & Sons, Hoboken, NJ, USA. pp. 261-284,
- 17. Baillot N, Girousse C, Allard V, Piquet-Pissaloux A, Le Gouis J. 2018. Different grain-filling rates explain grain-weight differences along the wheat ear. PLoS One 13: e0209597.
- 18. Balla K, Bed Z, Veisz O. 2007. Heat stress induced changes in the activity of antioxidant enzymes in wheat. Cereal Res Commun 35: 197-200.
- 19. Balla K, Bencze S, Janda T, Veisz O. 2009. Analysis of heat stress tolerance in winter wheat. Acta Agron Hunga 57: 437-444.
- 20. Bányai J, Karsai I, Balla K, Kiss T, Bed Z, Láng L. 2014. Heat stress response of wheat cultivars with different ecological adaptation. Cereal Res Commun 42: 413-425.
- 21. Barkley A, Tack J, Nalley LL, Bergtold J, Bowden R, Fritz A. 2014. Weather, disease, and wheat breeding effects on Kansas wheat varietal yields, 1985 to 2011. Agron J 106: 227-235.
- 22. Barnabás B, Jäger K, Fehér A. 2008. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ 31: 11-38.
- 23. Bavita A, Shashi B, Navtej SB. 2012. Nitric oxide alleviates oxidative damage induced by high temperature stress in wheat. Indian J Exp Biol 50: 372-378.
- 24. Bhat JA, Ali S, Salgotra RK, Mir ZA, Dutta S, Jadon V, Tyagi A, Mushtaq M, Jain N, Singh PK, Singh GP, Prabhu KV. 2016. Genomic selection in the era of next generation sequencing for complex traits in plant breeding. Front Genet 7: 221
- 25. Bhullar SS, Jenner CF. 1985. Differential responses to high temperatures of starch and nitrogen accumulation in the grain of four cultivars of wheat. Funct Plant Biol 12: 363-375.
- 26. Bhusal N, Sarial AK, Sharma P, Sareen S. 2017. Mapping QTLs for grain yield components in wheat under heat stress. PLoS One 12: e0189594.
- 27. Blumenthal C, Bekes F, Gras PW, Barlow EWR, Wrigley CW. 1995. Identification of wheat genotypes tolerant to the effects of heat stress on grain quality. Cereal Chem 72: 539-544.
- 28. Blumenthal CS, Batey IL, Bekes F, Wrigley CW, Barlow EWR. 1990. Gliadin genes contain heat-shock elements: possible relevance to heat-induced changes in grain quality. J Cereal Sci 11: 185-188.
- 29. Blumenthal CS, Bekes F, Batey IL, Wrigley CW, Moss HJ, Mares DJ, Barlow EWR. 1991. Interpretation of grain quality results from wheat variety trials with reference to high temperature stress. Aust J Agric Res 42: 325-334.
- 30. Bokshi AI, Thistlethwaite RJ, Chaplin ED, Kirii E, Trethowan RM, Tan DK. 2022. Physiological traits for evaluating heat-tolerance of Australian spring wheat cultivars at elevated CO2. J Agron Crop Sci 208: 178-196.
- 31. Bourgine B, Guihur A. 2021. Heat shock signaling in land plants: from plasma membrane sensing to the transcription of small heat shock proteins. Front Plant Sci 12: 710801.
- 32. Cairns JE, Prasanna BM. 2018. Developing and deploying climate-resilient maize varieties in the developing world. Curr Opin Plant Biol 45: 226-230.
- 33. Castro M, Peterson CJ, Rizza MD, Dellavalle PD, Vázquez D, Ibanez V, Ross A. In: Buck HT, Nisi JE, Salomón N. 2007. (Eds). Influence of heat stress on wheat grain characteristics and protein molecular weight distribution. . Wheat production in stressed environment. Developments in Plant Breeding. Springer, Dordrecht. pp. 365-371,
- 34. Caverzan A, Casassola A, Brammer SP. 2016. Antioxidant responses of wheat plants under stress. Genet Mol Biol 39: 1-6.
- 35. Chakraborty U, Pradhan D. 2011. High temperature-induced oxidative stress in Lens culinaris, role of antioxidants and amelioration of stress by chemical pre-treatments. J Plant Interact 6: 43-52.
- 36. Chen Z, Ke W, He F, Chai L, Cheng X, Xu H, Wang X, Du D, Zhao Y, Chen X, Xing J, Xin M, Guo W, Hu Z, Su Z, Liu J, Peng H, Yao Y, Sun Q, Ni Z. 2022. A single nucleotide deletion in the third exon of FT-D1increases the spikelet number and delays heading date in wheat (Triticum aestivum L.). Plant Biotechnol J 20: 920-933.
- 37. Cheng Z, Luan Y, Meng J, Sun J, Tao J, Zhao D. 2021. WRKY transcription factor response to high-temperature stress. Plants 10: 2211
- 38. Chiotelli E, Le Meste M. 2002. Effect of small and large wheat starch granules on thermomechanical behavior of starch. Cereal Chem 79: 286-293.
- 39. Cho C, Jeong H, Kim Y, Park J, Kim KH, Kim KM, Kang CS, Ko JM, Shon J. 2022. Growth and quality characteristics of Korean bread wheat in response to elevated temperature during their growing season. Korean J Crop Sci 67: 234-241.
- 40. Chunduri V, Kaur A, Kaur S, Kumar A, Sharma S, Sharma N, Singh P, Kapoor P, Kaur S, Kumari A, Roy J, Kaur J, Garg M. 2021. Gene expression and proteomics studies suggest an involvement of multiple pathways under day and day-night combined heat stresses during grain filling in wheat. Front Plant Sci 12: 660446.
- 41. Ciaffi M, Tozzi L, Borghi B, Corbellini M, Lafiandra D. 1996. Effect of heat shock during grain filling on the gluten protein composition of bread wheat. J Cereal Sci 24: 91-100.
- 42. Cossani CM, Reynolds MP. 2012. Physiological traits for improving heat tolerance in wheat. Plant Physiol 160: 1710-1718.
- 43. Crafts-Brandner SJ, Salvucci ME. 2000. Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2. PNAS 97: 13430-13435.
- 44. Cui X. 2020. Beyond yield response: weather shocks and crop abandonment. J Assoc Environ Reso 7: 901-932.
- 45. Daniel C, Triboi E. 2000. Effects of temperature and nitrogen nutrition on the grain composition of winter wheat: Effects on gliadin content and composition. J Cereal Sci 32: 45-56.
- 46. Darko E, Vegh B, Khalil R, Marcek T, Szalai G, Pal M, Janda T. 2019. Metabolic responses of wheat seedlings to osmotic stress induced by various osmolytes under iso-osmotic conditions. PLoS One 14: e0226151.
- 47. Davidson JF, Schiestl RH. 2001. Mitochondrial respiratory electron carriers are involved in oxidative stress during heat stress in Saccharomyces cerevisiae. Mol Cell Biol 21: 8483-8489.
- 48. Debbarma J, Sarki YN, Saikia B, Boruah HPD, Singha DL, Chikkaputtaiah C. 2019. Ethylene response factor (ERF) family proteins in abiotic stresses and CRISPR-Cas9 genome editing of ERFs for multiple abiotic stress tolerance in crop plants: A review. Mol Biotechnol 61: 153-172.
- 49. Dias AS, Bagulho AS, Lidon FC. 2008. Ultrastructure and biochemical traits of bread and durum wheat grains under heat stress. Braz J Plant Physiol 20: 323-333.
- 50. Dias AS, Lidon FC. 2009. Evaluation of grain filling rate and duration in bread and durum wheat, under heat stress after anthesis. J Agron Crop Sci 195: 137-147.
- 51. Djanaguiraman M, Boyle DL, Welti R, Jagadish SVK, Prasad PVV. 2018. Decreased photosynthetic rate under high temperature in wheat is due to lipid desaturation, oxidation, acylation, and damage of organelles. BMC Plant Biol 18: 55
- 52. Djanaguiraman M, Narayanan S, Erdayani E, Prasad PVV. 2020. Effects of high temperature stress during anthesis and grain filling periods on photosynthesis, lipids and grain yield in wheat. BMC Plant Biol 20: 268
- 53. Dupont FM, Altenbach SB. 2003. Molecular and biochemical impacts of environmental factors on wheat grain development and protein synthesis. J Cereal Sci 38: 133-146.
- 54. DuPont FM, Hurkman WJ, Vensel WH, Chan R, Lopez R, Tanaka CK, Altenbach SB. 2006. Differential accumulation of sulfur-rich and sulfur-poor wheat flour proteins is affected by temperature and mineral nutrition during grain development. J Cereal Sci 44: 101-112.
- 55. Dwivedi SK, Kumar G, Basu S, Kumar S, Rao KK, Choudhary AK. 2018. Physiological and molecular aspects of heat tolerance in wheat. SABRAO J Breed Genet 50: 192-216.
- 56. Dwivedi SK, Soni SK, Mishra JS, Koley TK, Kumar S. 2024. Assessment of terminal heat tolerance ability of wheat genotypes based on chemometric analysis and agro-physiological traits. Acta Physiol Plant 46: 48
- 57. El-Esawi MA, Al-Ghamdi AA, Ali HM, Ahmad M. 2019. Overexpression of AtWRKY30 transcription factor enhances heat and drought stress tolerance in wheat (Triticum aestivum L.). Genes (Basel) 10: 163
- 58. Erdayani E, Nagarajan R, Grant NP, Gill KS. 2020. Genome-wide analysis of the HSP101/CLPB gene family for heat tolerance in hexaploid wheat. Sci Rep 10: 3948
- 59. Essemine J, Ammar S, Bouzid S. 2010. Impact of heat stress on germination and growth in higher plants: Physiological, biochemical and molecular repercussions and mechanisms of defence. J Biol Sci 10: 565-572.
- 60. FAO.2019. FAOSTAT. http://www.fao.org/faostat/en/#data/FBS.
- 61. Farhad M, Kumar U, Tomar V, Bhati PK, Krishnan J N, Kishowar EM, Barek V, Brestic M, Hossain A. 2023. Heat stress in wheat: A global challenge to feed billions in the current era of the changing climate. Front Sustain Food Syst 7: 1203721.
- 62. Farooq M, Bramley H, Palta JA, Siddique KHM. 2011. Heat stress in wheat during reproductive and grain-filling phases. Crit Rev Plant Sci 30: 491-507.
- 63. Feng B, Liu P, Li G, Dong ST, Wang FH, Kong LA, Zhang JW. 2014. Effect of heat stress on the photosynthetic characteristics in flag leaves at the grain-filling stage of different heat-resistant winter wheat varieties. J Agron Crop Sci 200: 143-155.
- 64. Fernie E, Tan DKY, Liu SY, Ullah N, Khoddami A. 2022. Post-anthesis heat influences grain yield, physical and nutritional quality in wheat: A review. Agriculture 12: 886
- 65. Fischer RA, Rees D, Sayre KD, Lu ZM, Condon AG, Saavedra AL. 1998. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci 38: 1467-1475.
- 66. Fleitas MC, Mondal S, Gerard GS, Hernández-Espinosa N, Singh RP, Crossa J, Guzmán C. 2020. Identification of CIMMYT spring bread wheat germplasm maintaining superior grain yield and quality under heat-stress. J Cereal Sci 93: 102981.
- 67. Fokar M, Blum A, Nguyen HT. 1998. Heat tolerance in spring wheat. Ⅱ. Grain filling. Euphytica 104: 9-15.
- 68. Fu YB. 2015. Understanding crop genetic diversity under modern plant breeding. Theor Appl Genet 128: 2131-2142.
- 69. Gammans M, Mérel P, Ortiz-Bobea A. 2017. Negative impacts of climate change on cereal yields: statistical evidence from France. Environ Res Lett 12: 054007.
- 70. Gill SS, Tuteja N. 2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48: 909-930.
- 71. Gounaris K, Brain ARR, Quinn PJ, Williams WP. 1984. Structural reorganisation of chloroplast thylakoid membranes in response to heat-stress. BBA-Bioenergetics 766: 198-208.
- 72. Goyal M, Asthir B. 2010. Polyamine catabolism influences antioxidative defense mechanism in shoots and roots of five wheat genotypes under high temperature stress. Plant Growth Regul 60: 13-25.
- 73. Grant RF, Kimball BA, Conley MM, White JW, Wall GW, Ottman MJ. 2011. Controlled warming effects on wheat growth and yield: field measurements and modeling. Agron J 103: 1742-1754.
- 74. Grigorova B, Vassileva V, Klimchuk D, Vaseva I, Demirevska K, Feller U. 2012. Drought, high temperature, and their combination affect ultrastructure of chloroplasts and mitochondria in wheat (Triticum aestivum L.) leaves. J Plant Interact 7: 204-213.
- 75. Gudi S, Jain M, Singh S, Kaur S, Srivastava P, Mavi GS, Chhuneja P, Sohu V, Safhi FA, El-Moneim D A, Sharma A. 2024. Stress adaptive plasticity from Aegilops tauschii introgression lines improves drought and heat stress tolerance in bread wheat (Triticum aestivum L.). PeerJ 12: e17528.
- 76. Guérin C, Dupuits C, Mouzeyar S, Roche J. 2022. Insights into four NAC transcription factors involved in grain development and in response to moderate heat in the Triticeae Tribe. Int J Mol Sci 23: 11672
- 77. Guo W, Zhang J, Zhang N, Xin M, Peng H, Hu Z, Ni Z, Du J. 2015. The wheat NAC transcription factor TaNAC2L is regulated at the transcriptional and post-translational levels and promotes heat stress tolerance in transgenic Arabidopsis. PLoS One 10: e0135667.
- 78. Guo XL, Yuan SN, Zhang HN, Zhang YY, Zhang YJ, Wang GY, Li YQ, Li GL. 2020. Heat-response patterns of the heat shock transcription factor family in advanced development stages of wheat (Triticum aestivum L.) and thermotolerance-regulation by TaHsfA2-10. BMC Plant Biol 20: 364
- 79. Gupta A, Hua L, Zhang Z, Yang B, Li W. 2023. CRISPR-induced miRNA156-recognition element mutations in TaSPL13 improve multiple agronomic traits in wheat. Plant Biotechnol J 21: 536-548.
- 80. Gupta NK, Agarwal S, Agarwal VP, Nathawat NS, Gupta S, Singh G. 2013. Effect of short-term heat stress on growth, physiology and antioxidative defence system in wheat seedlings. Acta Physiol Plant 35: 1837-1842.
- 81. Hafeez MB, Zahra N, Kausar A, Li J, Rehman A, Farooq M. 2023. Influence of heat stress during grain development on the wheat grain yield, quality, and composition. J Soil Sci Plant Nut 23: 2955-2969.
- 82. Haider S, Raza A, Iqbal J, Shaukat M, Mahmood T. 2022. Analyzing the regulatory role of heat shock transcription factors in plant heat stress tolerance: a brief appraisal. Mol Biol Rep 49: 5771-5785.
- 83. Hasanuzzaman M, Nahar K, Alam MM, Roychowdhury R, Fujita M. 2013. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int J Mol Sci 14: 9643-9684.
- 84. Haslbeck M, Vierling E. 2015. A first line of stress defense: small heat shock proteins and their function in protein homeostasis. J Mol Biol 427: 1537-1548.
- 85. Hawker JS, Jenner CF. 1993. High temperature affects the activity of enzymes in the committed pathway of starch synthesis in developing wheat endosperm. Funct Plant Biol 20: 197-209.
- 86. Heidari P, Rezaee S, Hosseini Pouya HS, Mora-Poblete F. 2024. Insights into the Heat Shock Protein 70 (Hsp70) family in Camelina sativa and its roles in response to salt Stress. Plants 13: 3410
- 87. Hennessy K, Fawcett R, Kirono D, Mpelasoka F, Jones D, Bathols JM, Whetton P, Stafford Smith M, Howden M, Mitchell C. 2008. An assessment of the impact of climate change on the nature and frequency of exceptional climatic events. Bureau of Meteorology and CSIRO, Canberra, Melbourne..
- 88. Horváth I, Multhoff G, Sonnleitner A, Vigh L. 2008. Membrane-associated stress proteins: more than simply chaperones. BBA-Biomembranes 1778: 1653-1664.
- 89. Hossain A, Teixeira da Silva JA. 2012. Phenology, growth and yield of three wheat (Triticum aestivum L.) varieties as affected by high temperature stress. Not Sci Biol 4: 97-109.
- 90. Huang B, Rachmilevitch S, Xu J. 2012. Root carbon and protein metabolism associated with heat tolerance. J Exp Bot 63: 3455-3465.
- 91. Hurkman WJ, McCue KF, Altenbach SB, Korn A, Tanaka CK, Kothari KM, Johnson EL, Bechtel DB, Wilson JD, Anderson OD, DuPont FM. 2003. Effect of temperature on expression of genesencoding enzymes for starch biosynthesis in developing wheat endosperm. Plant Sci 164: 873-881.
- 92. Hurkman WJ, Vensel WH, Tanaka CK, Whitehand L, Altenbach SB. 2009. Effect of high temperature on albumin and globulin accumulation in the endosperm proteome of the developing wheat grain. J Cereal Sci 49: 12-23.
- 93. Hurkman WJ, Wood DF. 2011. High temperature during grain fill alters the morphology of protein and starch deposits in the starchy endosperm cells of developing wheat (Triticum aestivum L.) grain. J Agric Food Chem 59: 4938-4946.
- 94. Hussain B, Lucas SJ, Budak H. 2018. CRISPR/Cas9 in plants: at play in the genome and at work for crop improvement. Brief Funct Genomics 17: 319-328.
- 95. Janni M, Gulli M, Maestri E, Marmiroli M, Valliyodan B, Nguyen HT, Marmiroli N. 2020. Molecular and genetic bases of heat stress responses in crop plants and breeding for increased resilience and productivity. J Exp Bot 71: 3780-3802.
- 96. Jenner CF, Ugalde TD, Aspinall D. 1991. The physiology of starch and protein deposition in the endosperm of wheat. Funct Plant Biol 18: 211-226.
- 97. Jenner CF. 1994. Starch synthesis in the kernel of wheat under high temperature conditions. Funct Plant Biol 21: 791-806.
- 98. Jeon JS, Ryoo N, Hahn TR, Walia H, Nakamura Y. 2010. Starch biosynthesis in cereal endosperm. Plant Physiol Biochem 48: 383-392.
- 99. Jing J, Guo S, Li Y, Li W. 2020. The alleviating effect of exogenous polyamines on heat stress susceptibility of different heat resistant wheat (Triticum aestivum L.) varieties. Sci Rep 10: 7467
- 100. Jouanin A, Gilissen L, Schaart JG, Leigh FJ, Cockram J, Wallington EJ, Boyd LA, van den Broeck HC, van der Meer IM, America AHP, Visser RGF, Smulders MJM. 2020. CRISPR/Cas9 gene editing of gluten in wheat to reduce gluten content and exposure-Reviewing methods to screen for coeliac safety. Front Nutr 7: 51
- 101. Juliana P, Montesinos-López OA, Crossa J, Mondal S, González Pérez L, Poland J, Huerta-Espino J, Crespo-Herrera L, Govindan V, Dreisigacker S, Shrestha S, Pérez-Rodríguez P, Pinto Espinosa F, Singh RP. 2019. Integrating genomic-enabled prediction and high-throughput phenotyping in breeding for climate-resilient bread wheat. Theor Appl Genet 132: 177-194.
- 102. Kaushal N, Bhandari K, Siddique KHM, Nayyar H, Tejada Moral M. 2016. Food crops face rising temperatures: An overview of responses, adaptive mechanisms, and approaches to improve heat tolerance. Cogent Food Agric 2: 1134380.
- 103. Keeling PL, Bacon PJ, Holt DC. 1993. Elevated temperature reduces starch deposition in wheat endosperm by reducing the activity of soluble starch synthase. Planta 191: 342-348.
- 104. Khan A, Ahmad M, Ahmed M, Iftikhar Hussain M. 2020. Rising atmospheric temperature impact on wheat and thermotolerance strategies. Plants 10: 43
- 105. Khanna-Chopra R, Chauhan S. 2015. Wheat cultivars differing in heat tolerance show a differential response to oxidative stress during monocarpic senescence under high temperature stress. Protoplasma 252: 1241-1251.
- 106. Khanzada A, Wang X, Malko MM, Wu Y, Samo A, Dong J. 2024. Response of the persistence of heat stress tolerance in winter wheat seedling to heat priming at early growth stages. Plant Stress 11: 100323.
- 107. Kim D, Alptekin B, Budak H. 2018. CRISPR/Cas9 genome editing in wheat. Funct Integr Genomic 18: 31-41.
- 108. Kocsy G, Szalai G, Galiba G. 2002. Effect of heat stress on glutathione biosynthesis in wheat. Acta Biol Szeged 46: 71-72.
- 109. Koga S, Böcker U, Moldestad A, Tosi P, Shewry PR, Mosleth EF, Uhlen AK. 2015. Influence of temperature on the composition and polymerization of gluten proteins during grain filling in spring wheat (Triticum aestivum L.). J Cereal Sci 65: 1-8.
- 110. Kosova K, Vitamvas P, Prasil IT, Renaut J. 2011. Plant proteome changes under abiotic stress-contribution of proteomics studies to understanding plant stress response. J Proteomics 74: 1301-1322.
- 111. Kumar A, Sharma S, Chunduri V, Kaur A, Kaur S, Malhotra N, Kumar A, Kapoor P, Kumari A, Kaur J, Sonah H, Garg M. 2020. Genome-wide identification and characterization of heat shock protein family reveals role in development and stress conditions in Triticum aestivum L. Sci Rep 10: 7858
- 112. Kumar R, Tripathi G, Goyal I, Sharma J, Tiwari R, Shimphrui R, Sarkar NK, Grover A. 2023. Insights into genomic variations in rice Hsp100 genes across diverse rice accessions. Planta 257: 91
- 113. Kumar RR, Goswami S, Sharma SK, Singh K, Gadpayle KA, Kumar N, Ral GK, Singh M, Ral RD. 2012. Protection against heat stress in wheat involves change in cell membrane stability, antioxidant enzymes, osmolyte, H2O2 and transcript of heat shock protein. Int J Plant Physiol Biochem 4: 83-91.
- 114. Kumar RR, Goswami S, Singh K, Dubey K, Rai GK, Singh B, Singh S, Grover M, Mishra D, Kumar S, Bakshi S, Rai A, Pathak H, Chinnusamy V, Praveen S. 2018. Characterization of novel heat-responsive transcription factor (TaHSFA6e) gene involved in regulation of heat shock proteins (HSPs)-A key member of heat stress-tolerance network of wheat. J Biotechnol 279: 1-12.
- 115. Kumar RR, Tasleem M, Jain M, Ahuja S, Goswami S, Bakshi S, Jambhulkar S, Singh SD, Singh GP, Pathak H, Viswanathan C, Praveen S. 2019. Nitric oxide triggered defense network in wheat: Augmenting tolerance and grain-quality related traits under heat-induced oxidative damage. Environ Exp Bot 158: 189-204.
- 116. Kumar S, Singh R, Nayyar H. 2013. α-Tocopherol application modulates the response of wheat (Triticum aestivum L.) seedlings to elevated temperatures by mitigation of stress injury and enhancement of antioxidants. J Plant Growth Regul 32: 307-314.
- 117. Kumari M, Pudake RN, Singh VP, Joshi AK. 2013. Association of staygreen trait with canopy temperature depression and yield traits under terminal heat stress in wheat (Triticum aestivum L.). Euphytica 190: 87-97.
- 118. Lal MK, Tiwari RK, Gahlaut V, Mangal V, Kumar A, Singh MP, Paul V, Kumar S, Singh B, Zinta G. 2022. Physiological and molecular insights on wheat responses to heat stress. Plant Cell Rep 41: 501-518.
- 119. Lamaoui M, Jemo M, Datla R, Bekkaoui F. 2018. Heat and drought stresses in crops and approaches for their mitigation. Front Chem 6: 26
- 120. Lamba K, Kumar M, Singh V, Chaudhary L, Sharma R, Yashveer S, Dalal MS. 2023. Heat stress tolerance indices for identification of the heat tolerant wheat genotypes. Sci Rep-UK 13: 10842
- 121. Langridge P, Reynolds MP. 2015. Genomic tools to assist breeding for drought tolerance. Curr Opin Biotechnol 32: 130-135.
- 122. Larkin DL, Holder AL, Mason RE, Moon DE, Brown-Guedira G, Price PP, Harrison SA, Dong Y. 2020. Genome-wide analysis and prediction of Fusarium head blight resistance in soft red winter wheat. Crop Sci 60: 2882-2900.
- 123. Latif S, Wang L, Khan J, Ali Z, Sehgal SK, Babar MA, Wang J, Quraishi UM. 2020. Deciphering the role of stay-green trait to mitigate terminal heat stress in bread wheat. Agronomy 10: 1001
- 124. Levitt J. 1980. Responses of plant to environmental stress: Water, radiation, salt and other stresses. Academic Press, New York. p. 365..
- 125. Li H, Liu H, Hao C, Li T, Liu Y, Wang X, Yang Y, Zheng J, Zhang X. 2023. The auxin response factor TaARF15-A1 negatively regulates senescence in common wheat (Triticum aestivum L.). Plant Physiol 191: 1254-1271.
- 126. Li R, Hou L, Zhang A, Lu Y, Song W, Tadesse W, Wang X, Liu M, Zheng W, Xu S. 2018. Heat stress in filling stage confers distinct effect on starch granules formation in different thermotolerant wheat accessions. Pak J Bot 50: 913-920.
- 127. Lin Q, Zong Y, Xue C, Wang S, Jin S, Zhu Z, Wang Y, Anzalone AV, Raguram A, Doman JL, Liu DR, Gao C. 2020. Prime genome editing in rice and wheat. Nat Biotechnol 38: 582-585.
- 128. Liu B, Asseng S, Wang A, Wang S, Tang L, Cao W, Zhu Y, Liu L. 2017. Modelling the effects of post-heading heat stress on biomass growth of winter wheat. Agr Forest Meteorol 247: 476-490.
- 129. Liu P, Guo W, Jiang Z, Pu H, Feng C, Zhu X, Peng Y, Kuang A, Little CR. 2011. Effects of high temperature after anthesis on starch granules in grains of wheat (Triticum aestivum L.). J Agric Sci 149: 159-169.
- 130. Liu X, Huang B. 2000. Heat stress injury in relation to membrane lipid peroxidation in creeping bentgrass. Crop Sci 40: 503-510.
- 131. Liu X, Zhou D, Dai C, Zhu Y, Zhu M, Ding J, Zhu X, Zhou G, Guo W, Li C. 2023. Difference in starch structure and physicochemical properties between waxy wheat and non-waxy wheat subjected to temporary heat stress during grain filling. Agronomy 13: 2067
- 132. Lu H, Wang C, Guo T, Xie Y, Feng W, Li S. 2014. Starch composition and its granules distribution in wheat grains in relation to post-anthesis high temperature and drought stress treatments. Starch - Stärke 66: 419-428.
- 133. Lu L, Liu H, Wu Y, Yan G. 2022. Wheat genotypes tolerant to heat at seedling stage tend to be also tolerant at adult stage: The possibility of early selection for heat tolerance breeding. Crop J 10: 1006-1013.
- 134. Majoul T, Bancel E, Triboï E, Ben Hamida J, Branlard G. 2003. Proteomic analysis of the effect of heat stress on hexaploid wheat grain: Characterization of heat-responsive proteins from total endosperm. Proteomics 3: 175-183.
- 135. Majoul T, Bancel E, Triboi E, Ben Hamida J, Branlard G. 2004. Proteomic analysis of the effect of heat stress on hexaploid wheat grain: characterization of heat-responsive proteins from non-prolamins fraction. Proteomics 4: 505-513.
- 136. Martínez-Goñi XS, Miranda-Apodaca J, Pérez-López U. 2024. Enhanced photosynthesis, transpiration regulation, water use-efficiency and growth in buckwheat outperforms wheat response to high [CO2], high temperature and drought. Environ Exp Bot 222: 105756.
- 137. Mathur S, Agrawal D, Jajoo A. 2014. Photosynthesis: Response to high temperature stress. J Photoch Photobio B 137: 116-126.
- 138. Mathur S, Jajoo A, Mehta P, Bharti S. 2011. Analysis of elevated temperature-induced inhibition of photosystem Ⅱ using chlorophyll a fluorescence induction kinetics in wheat leaves (Triticum aestivum). Plant Biol (Stuttg) 13: 1-6.
- 139. McClung CR, Davis SJ. 2010. Ambient thermometers in plants: from physiological outputs towards mechanisms of thermal sensing. Curr Biol 20: R1086-1092.
- 140. Mirosavljević M, Mikić S, Kondić-Špika A, Župunski V, Zhou R, Abdelhakim L, Ottosen CO. 2021b. The effect of heat stress on some main spike traits in 12 wheat cultivars at anthesis and mid-grain filling stage. Plant Soil Environ 67: 71-76.
- 141. Mirosavljević M, Mikić S, Župunski V, Abdelhakim L, Trkulja D, Zhou R, Kondić Špika A, Ottosen CO. 2024. Effects of heat stress during anthesis and grain filling stages on some physiological and agronomic traits in diverse wheat genotypes. Plants 13: 2083
- 142. Mirosavljević M, Mikić S, Župunski V, Kondić Špika A, Trkulja D, Ottosen CO, Zhou R, Abdelhakim L. 2021a. Effects of high temperature during anthesis and grain filling on physiological characteristics of winter wheat cultivars. J Agron Crop Sci 207: 823-832.
- 143. Mishra R, Joshi RK, Zhao K. 2020. Base editing in crops: current advances, limitations and future implications. Plant Biotechnol J 18: 20-31.
- 144. Mittler R, Vanderauwera S, Gollery M, Van Breusegem F. 2004. Reactive oxygen gene network of plants. Trends Plant Sci 9: 490-498.
- 145. Mittler R, Zandalinas SI, Fichman Y, Van Breusegem F. 2022. Reactive oxygen species signalling in plant stress responses. Nat Rev Mol Cell Biol 23: 663-679.
- 146. Mittler R. 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 7: 405-410.
- 147. Mohi-Ud-Din M, Siddiqui N, Rohman M, Jagadish SK, Ahmed JU, Hassan MM, Hossain A, Islam T. 2021. Physiological and biochemical dissection reveals a trade-off between antioxidant capacity and heat tolerance in bread wheat (Triticum aestivum L.). Antioxidants 10: 351
- 148. Molero G, Coombes B, Joynson R, Pinto F, Piñera-Chávez FJ, Rivera-Amado C, Hall A, Reynolds MP. 2023. Exotic alleles contribute to heat tolerance in wheat under field conditions. Commun Biol 6: 21
- 149. Mondal S, Sallam A, Sehgal D, Sukumaran S, Farhad M, Navaneetha Krishnan J, Kumar U, Biswal A. 2021. Advances in breeding for abiotic stress tolerance in wheat. pp 71-103. In: Kole C. (Ed) Genomic designing for abiotic stress resistant cereal crops. Springer, Cham:
- 150. Morran S, Eini O, Pyvovarenko T, Parent B, Singh R, Ismagul A, Eliby S, Shirley N, Langridge P, Lopato S. 2011. Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors. Plant Biotechnol J 9: 230-249.
- 151. Mullineaux PM, Baker NR. 2010. Oxidative stress: antagonistic signaling for acclimation or cell death? Plant physiol 154: 521-525.
- 152. Nahar K, Ahamed KU, Fujita M. 2010. Phenological variation and its relation with yield in several wheat (Triticum aestivum L.) cultivars under normal and late sowing mediated heat stress condition. Not Sci Bio 2: 51-56.
- 153. Nawaz A, Farooq M, Cheema SA, Wahid A. 2013. Differential response of wheat cultivars to terminal heat stress. Int J Agric Biol 15: 1354-1358.
- 154. Nicolas ME, Lambers H, Simpson RJ, Dalling MJ. 1985. Effect of drought on metabolism and partitioning of carbon in two wheat varieties differing in drought-tolerance. Ann Bot 55: 727-742.
- 155. Niu X, Luo T, Zhao H, Su Y, Ji W, Li H. 2020. Identification of wheat DREB genes and functional characterization of TaDREB3 in response to abiotic stresses. Gene 740: 144514.
- 156. Noctor G, Foyer CH. 1998. Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Biol 49: 249-279.
- 157. Paliwal R, Röder MS, Kumar U, Srivastava JP, Joshi AK. 2012. QTL mapping of terminal heat tolerance in hexaploid wheat (T. aestivum L.). Theor Appl Genet 125: 561-575.
- 158. Pandey B, Kaur A, Gupta OP, Sharma I, Sharma P. 2015. Identification of HSP20 gene family in wheat and barley and their differential expression profiling under heat stress. Appl Biochem Biotech 175: 2427-2446.
- 159. Park S-H, Wilson JD, Seabourn BW. 2009. Starch granule size distribution of hard red winter and hard red spring wheat: Its effects on mixing and breadmaking quality. J Cereal Sci 49: 98-105.
- 160. Patidar A, Yadav MC, Kumari J, Tiwari S, Chawla G, Paul V. 2023. Identification of climate-smart bread wheat germplasm lines with enhanced adaptation to global warming. Plants 12: 2851
- 161. Paulsen GM. In: Boote KJ, Bennett JM, Sinclair TR, Paulsen GM. 1994. (Eds). High temperature responses of crop plants. . Physiology and determination of crop yield. ASA, CSSA, and SSSA, Madison. pp. 365-389,
- 162. Peng J, Sun D, Peng Y, Nevo E. 2013. Gene discovery in Triticum dicoccoides, the direct progenitor of cultivated wheats. Cereal Res Commun 41: 1-22.
- 163. Peterson DG, Fulcher R. 2001. Variation in Minnesota HRS wheats: starch granule size distribution. Food Res Int 34: 357-363.
- 164. Pinto RS, Lopes MS, Collins NC, Reynolds MP. 2016. Modelling and genetic dissection of staygreen under heat stress. Theor Appl Genet 129: 2055-2074.
- 165. Poudel PB, Poudel MR. 2020. Heat stress effects and tolerance in wheat: A review. J Biol Today'. s World 9: 217.
- 166. Pradhan GP, Prasad PV. 2015. Evaluation of wheat chromosome translocation lines for high temperature stress tolerance at grain filling stage. PLoS One 10: e0116620.
- 167. Prakash P, Sharma-Natu P, Ghildiyal M. 2004. Effect of different temperature on starch synthase activity in excised grains of wheat cultivars. Indian J Exp Biol 42: 227-230.
- 168. Prasad PVV, Staggenborg SA, Ristic Z. In: Ahuja LR, Reddy VR, Saseendran SA, Yu Q. 2008. (Eds). Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. . Response of crops to limited water: Understanding and modeling water stress effects on plant growth processes. ASA, CSSA, Madison. pp. 301-355,
- 169. Qaseem MF, Qureshi R, Shaheen H. 2019. Effects of pre-anthesis drought, heat and their combination on the growth, yield and physiology of diverse wheat (Triticum aestivum L.) genotypes varying in sensitivity to heat and drought stress. Sci Rep 9: 6955
- 170. Raffan S, Sparks C, Huttly A, Hyde L, Martignago D, Mead A, Hanley SJ, Wilkinson PA, Barker G, Edwards KJ, Curtis TY, Usher S, Kosik O, Halford NG. 2021. Wheat with greatly reduced accumulation of free asparagine in the grain, produced by CRISPR/Cas9 editing of asparagine synthetase gene TaASN2. Plant Biotechnol J 19: 1602-1613.
- 171. Rahman MA, Chikushi J, Yoshida S, Karim A. 2009. Growth and yield components of wheat genotypes exposed to high temperature stress under control environment. Bangladesh J Agric Res 34: 360-372.
- 172. Rakszegi M, Lovegrove A, Balla K, Láng L, Bedő Z, Veisz O, Shewry PR. 2014. Effect of heat and drought stress on the structure and composition of arabinoxylan and β-glucan in wheat grain. Carbohyd Polym 102: 557-565.
- 173. Rangan P, Furtado A, Henry R. 2019. Differential response of wheat genotypes to heat stress during grain filling. Exp Agr 55: 818-827.
- 174. Reynolds M, Tattaris M, Cossani CM, Ellis M, Yamaguchi-Shinozaki K, Pierre CS. 2015. Exploring genetic resources to increase adaptation of wheat to climate change. In: Advances in wheat genetics: From genome to field. Proc 12th Int Wheat Genet Symp, Springer, Berlin/Heidelberg, Germany:
- 175. Reynolds MP, Balota M, Delgado MIB, Amani I, Fischer RA. 1994. Physiological and morphological traits associated with spring wheat yield under hot, irrigated conditions. Funct Plant Biol 21: 717-730.
- 176. Reynolds MP, Gutiérrez-Rodrı́guez M, Larqué-Saavedra A. 2000. Photosynthesis of wheat in a warm, irrigated environment: I: Genetic diversity and crop productivity. Field Crops Res 66: 37-50.
- 177. Ristic Z, Bukovnik U, Prasad PVV. 2007. Correlation between heat stability of thylakoid membranes and loss of chlorophyll in winter wheat under heat stress. Crop Sci 47: 2067-2073.
- 178. Ristic Z, Momcilovic I, Bukovnik U, Prasad PV, Fu J, Deridder BP, Elthon TE, Mladenov N. 2009. Rubisco activase and wheat productivity under heat-stress conditions. J Exp Bot 60: 4003-4014.
- 179. Rosenow DT, Quisenberry JE, Wendt CW, Clark LE. 1983. Drought tolerant sorghum and cotton germplasm. Agr Water Manage 7: 207-222.
- 180. Roy C, Chattopadhyay T, Ranjan RD, Ul Hasan W, Kumar A, De N. 2021. Association of leaf chlorophyll content with the stay-green trait and grain yield in wheat grown under heat stress conditions. Czech J Genet Plant Breed 57: 140-148.
- 181. Royo C, Briceño-Félix GA. 2011. Spanish wheat pool. In: The world wheat book. Vol. 2: Lavoisier Publishing Inc., Paris: pp. 121-154.
- 182. Ruelland E, Zachowski A. 2010. How plants sense temperature. Environ Exp Bot 69: 225-232.
- 183. Salvucci ME, Crafts-Brandner SJ. 2004a. Relationship between the heat tolerance of photosynthesis and the thermal stability of rubisco activase in plants from contrasting thermal environments. Plant Physiol 134: 1460-1470.
- 184. Salvucci ME, Crafts-Brandner SJ. 2004b. Mechanism for deactivation of Rubisco under moderate heat stress. Physiol Plant 122: 513-519.
- 185. Salvucci ME, Crafts-Brandner SJ. 2004c. Inhibition of photosynthesis by heat stress: the activation state of Rubisco as a limiting factor in photosynthesis. Physiol Plantarum 120: 179-186.
- 186. Sarkar J, Chakraborty B, Chakraborty U. 2016. Temperature stress induced antioxidative and biochemical changes in wheat (Triticum aestivumL.) cultivars. J Plant Stress Physiol 2: 22-30.
- 187. Savicka M, Škute N. 2010. Effects of high temperature on malondialdehyde content, superoxide production and growth changes in wheat seedlings (Triticum aestivum L.). Ekologija 56: 26-33.
- 188. Scharf KD, Berberich T, Ebersberger I, Nover L. 2012. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. BBA-Gene Regul Mech 1819: 104-119.
- 189. Semenov MA, Shewry PR. 2011. Modelling predicts that heat stress, not drought, will increase vulnerability of wheat in Europe. Sci Rep 1: 66
- 190. Sharma D, Singh R, Tiwari R, Kumar R, Gupta VK. In: Hasanuzzaman M, Nahar K, Hossain MA. 2019. (Eds). Wheat responses and tolerance to terminal heat stress: A review. . Wheat production in changing environments: Responses, adaptation and tolerance. Springer, Singapore. pp. 149-173,
- 191. Sharma P, Jha AB, Dubey RS, Pessarakli M. 2012. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J Bot 2012: 217037.
- 192. Shirdelmoghanloo H, Lohraseb I, Rabie HS, Brien C, Parent B, Collins NC. 2016. Heat susceptibility of grain filling in wheat (Triticum aestivum L.) linked with rapid chlorophyll loss during a 3-day heat treatment. Acta Physiol Plant 38: 208
- 193. Singh MK, Shin Y, Ju S, Han S, Choe W, Yoon KS, Kim SS, Kang I. 2024. Heat shock response and heat shock proteins: Current understanding and future opportunities in human diseases. Int J Mol Sci 25: 4209
- 194. Singh N, Virdi AS, Katyal M, Kaur A, Kaur D, Ahlawat AK, Singh AM, Kumar Sharma R. 2021. Evaluation of heat stress through delayed sowing on physicochemical and functional characteristics of grains, whole meals and flours of India wheat. Food Chem 344: 128725.
- 195. Singha DL, Das D, Paswan RR, Chikkaputtaiah C, Kumar S. In: Sahu J, Vaishnav A, Sing HB. 2022. (Eds). Novel approaches and advanced molecular techniques for crop improvement. . Plant-microbe interactions. CRC Press, Boca Raton, FL, USA. pp. 1-27,
- 196. Skylas DJ, Cordwell SJ, Hains PG, Larsen MR, Basseal DJ, Walsh BJ, Blumenthal C, Rathmell W, Copeland L, Wrigley CW. 2002. Heat shock of wheat during grain filling: proteins associated with heat-tolerance. J Cereal Sci 35: 175-188.
- 197. Sofield I, Evans L, Cook M, Wardlaw I. 1977. Factors influencing the rate and duration of grain filling in wheat. Funct Plant Biol 4: 785-797.
- 198. Sofo A, Scopa A, Nuzzaci M, Vitti A. 2015. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int J Mol Sci 16: 13561-13578.
- 199. Soliman WS, Fujimori M, Tase K, Sugiyama S-i. 2011. Oxidative stress and physiological damage under prolonged heat stress in C3 grass Lolium perenne. Grassl Sci 57: 101-106.
- 200. Son JH, Kang C-S, Yoon YM, Choi CH, Kim KH, Kim KM, Park TI, Kang TG, Kang SW, Park CS, Cho SW. 2019. Effect of high temperature during grain maturation on flour properties and end-use quality in Korean wheat cultivars. Korean J Breed Sci 51: 20-33.
- 201. Song J, Carver BF, Powers C, Yan L, Klapste J, El-Kassaby YA, Chen C. 2017. Practical application of genomic selection in a doubled-haploid winter wheat breeding program. Mol Breed 37: 117
- 202. Stone PJ, Nicolas ME. 1995. Effect of timing of heat stress during grain filling on two wheat varieties differing in heat tolerance. I. Grain growth. Funct Plant Biol 22: 927-934.
- 203. Stone PJ, Nicolas ME. 1996. Effect of timing of heat stress during grain filling on two wheat varieties differing in heat tolerance. Ⅱ. Fractional protein accumulation. Funct Plant Biol 23: 739-749.
- 204. Stone PJ, Savin R, Wardlaw IF, Nicolas ME. 1995. The influence of recovery temperature on the effects of a brief heat shock on wheat. I. Grain growth. Funct Plant Biol 22: 945-954.
- 205. Streck NA. 2005. Climate change and agroecosystems: The effect of elevated atmospheric CO2 and temperature on crop growth, development, and yield. Cienc Rural 35: 730-740.
- 206. Tashiro T, Wardlaw IF. 1989. A comparison of the effect of high temperature on grain development in wheat and rice. Ann Bot 64: 59-65.
- 207. Tayade R, Nguyen T, Oh SA, Hwang YS, Yoon IS, Deshmuk R, Jung K-H, Park SK. 2018. Effective strategies for enhancing tolerance to high-temperature stress in rice during the reproductive and ripening stages. Plant Breed Biotech 6: 1-18.
- 208. Thomas H, Howarth CJ. 2000. Five ways to stay green. J Exp Bot 51: 329-337.
- 209. Tomás D, Rodrigues JC, Viegas W, Silva M. 2020. Assessment of high temperature effects on grain yield and composition in bread wheat commercial varieties. Agronomy 10: 499
- 210. Trethowan RM, Mujeeb-Kazi A. 2008. Novel germplasm resources for improving environmental stress tolerance of hexaploid wheat. Crop Sci 48: 1255-1265.
- 211. Tricker PJ, ElHabti A, Schmidt J, Fleury D. 2018. The physiological and genetic basis of combined drought and heat tolerance in wheat. J Exp Bot 69: 3195-3210.
- 212. Tuinstra MR, Grote EM, Goldsbrough PB, Ejeta G. 1997. Genetic analysis of post-flowering drought tolerance and components of grain development in Sorghum bicolor (L.) Moench. Mol Breeding 3: 439-448.
- 213. Ullah A, Nadeem F, Nawaz A, Siddique KHM, Farooq M. 2022. Heat stress effects on the reproductive physiology and yield of wheat. J Agron Crop Sci 208: 1-17.
- 214. Usman MG, Rafii M, Ismail M, Malek M, Latif MA, Oladosu Y. 2014. Heat shock proteins: Functions and response against heat stress in plants. Int J Sci Technol Res 3: 204-218.
- 215. Vignjevic M, Wang X, Olesen JE, Wollenweber B. 2015. Traits in spring wheat cultivars associated with yield loss caused by a heat stress episode after anthesis. J Agron Crop Sci 201: 32-48.
- 216. Vincent CD, Gregory PJ. 1989. Effects of temperature on the development and growth of winter wheat roots: Ⅰ. Controlled glasshouse studies of temperature, nitrogen and irradiance. Plant Soil 119: 87-97.
- 217. Viswanathan C, Khanna-Chopra R. 2001. Effect of heat stress on grain growth, starch synthesis and protein synthesis in grains of wheat (Triticum aestivum L.) varieties differing in grain weight stability. J Agron Crop Sci 186: 1-7.
- 218. Wang W, Pan Q, He F, Akhunova A, Chao S, Trick H, Akhunov E. 2018. Transgenerational CRISPR-Cas9 activity facilitates multiplex gene editing in allopolyploid wheat. Cris J 1: 65-74.
- 219. Wang W, Pan Q, Tian B, He F, Chen Y, Bai G, Akhunova A, Trick HN, Akhunov E. 2019. Gene editing of the wheat homologs of TONNEAU1-recruiting motif encoding gene affects grain shape and weight in wheat. Plant J 100: 251-264.
- 220. Wang W, Vinocur B, Shoseyov O, Altman A. 2004. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci 9: 244-252.
- 221. Wang X, Cai J, Liu F, Jin M, Yu H, Jiang D, Wollenweber B, Dai T, Cao W. 2012. Pre-anthesis high temperature acclimation alleviates the negative effects of post-anthesis heat stress on stem stored carbohydrates remobilization and grain starch accumulation in wheat. J Cereal Sci 55: 331-336.
- 222. Wang X, Dinler BS, Vignjevic M, Jacobsen S, Wollenweber B. 2015. Physiological and proteome studies of responses to heat stress during grain filling in contrasting wheat cultivars. Plant Sci 230: 33-50.
- 223. Wang X, Hou L, Lu Y, Wu B, Gong X, Liu M, Wang J, Sun Q, Vierling E, Xu S. 2018. Metabolic adaptation of wheat grain contributes to a stable filling rate under heat stress. J Exp Bot 69: 5531-5545.
- 224. Wang X, Xu Y, Hu Z, Xu C. 2018. Genomic selection methods for crop improvement: Current status and prospects. Crop J 6: 330-340.
- 225. Wang Y, Cheng X, Shan Q, Zhang Y, Liu J, Gao C, Qiu J. 2014. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat Biotechnol 32: 947-951.
- 226. Wardlaw IF, Blumenthal C, Larroque O, Wrigley CW. 2002. Contrasting effects of chronic heat stress and heat shock on kernel weight and flour quality in wheat. Funct Plant Biol 29: 25-34.
- 227. Wardlaw IF, Moncur L. 1995. The response of wheat to high temperature following anthesis. I. The rate and duration of kernel filling. Funct Plant Biol 22: 391-397.
- 228. Wardlaw IF, Sofield I, Cartwright PM. 1980. Factors limiting the rate of dry matter accumulation in the grain of wheat grown at high temperature. Funct Plant Biol 7: 387-400.
- 229. Wieser H, Koehler P, Scherf KA. 2023. Chemistry of wheat gluten proteins: Qualitative composition. Cereal Chem 100: 23-35.
- 230. Wolde GM, Mascher M, Schnurbusch T. 2019. Genetic modification of spikelet arrangement in wheat increases grain number without significantly affecting grain weight. Mol Genet Genomics 294: 457-468.
- 231. Xin M, Peng H, Ni Z, Yao Y, Hu Z, Sun Q. In: Hasanuzzaman M, Nahar K, Hossain MA. 2019. (Eds). Wheat responses and tolerance to high temperature. . Wheat production in changing environments: Responses, adaptation and tolerance. Springer, Singapore. pp. 139-147,
- 232. Xue GP, Drenth J, McIntyre CL. 2015. TaHsfA6f is a transcriptional activator that regulates a suite of heat stress protection genes in wheat (Triticum aestivum L.) including previously unknown Hsf targets. J Exp Bot 66: 1025-1039.
- 233. Yadav MR, Choudhary M, Singh J, Lal MK, Jha PK, Udawat P, Gupta NK, Rajput VD, Garg NK, Maheshwari C, Hassan M, Gupta S, Jatwa TK, Kuamr R, Yadav AK, Prasad PV. 2022. Impacts, tolerance, adaptation, and mitigation of heat stress on wheat under changing climates. Int J Mol Sci 23: 2838
- 234. Yadav SBK, Bhatt S. 2024. Heat stress and it's tolerance in wheat. Cogent Food Agric 10: 2413398.
- 235. Yamamoto Y, Aminaka R, Yoshioka M, Khatoon M, Komayama K, Takenaka D, Yamashita A, Nijo N, Inagawa K, Morita N, Sasaki T, Yamamoto Y. 2008. Quality control of photosystem Ⅱ: Impact of light and heat stresses. Photosynth Res 98: 589-608.
- 236. Yan SH, Yin Y, Li WY, Liang TB. 2008. Effect of high temperature during grain filling on starch accumulation, starch granule distribution, and activities of related enzymes in wheat grains. Acta Agronomica Sinica 34: 1092-1096.
- 237. Yang J, Sears R, Gill B, Paulsen G. 2002. Growth and senescence characteristics associated with tolerance of wheat-alien amphiploids to high temperature under controlled conditions. Euphytica 126: 185-193.
- 238. Yang J, Zhang J, Wang Z, Xu G, Zhu Q. 2004. Activities of key enzymes in sucrose-to-starch conversion in wheat grains subjected to water deficit during grain filling. Plant Physiol 135: 1621-1629.
- 239. Yang X, Tian Z, Sun L, Chen B, Tubiello FN, Xu Y. 2017. The impacts of increased heat stress events on wheat yield under climate change in China. Climatic Change 140: 605-620.
- 240. Yin X, Guo W, Spiertz JH. 2009. A quantitative approach to characterize sink-source relationships during grain filling in contrasting wheat genotypes. Field Crops Res 114: 119-126.
- 241. Yu Z, Yunusbaev U, Fritz A, Tilley M, Akhunova A, Trick H, Akhunov E. 2024. CRISPR-based editing of the omega- and gamma-gliadin gene clusters reduces wheat immunoreactivity without affecting grain protein quality. Plant Biotechnol J 22: 892-903.
- 242. Zang X, Geng X, He K, Wang F, Tian X, Xin M, Yao Y, Hu Z, Ni Z, Sun Q, Peng H. 2018. Overexpression of the wheat (Triticum aestivum L.) TaPEPKR2 gene enhances heat and dehydration tolerance in both wheat and Arabidopsis. Front Plant Sci 9: 1710
- 243. Zang X, Geng X, Wang F, Liu Z, Zhang L, Zhao Y, Tian X, Ni Z, Yao Y, Xin M, Hu Z, Sun Q, Peng H. 2017. Overexpression of wheat ferritin gene TaFER-5B enhances tolerance to heat stress and other abiotic stresses associated with the ROS scavenging. BMC Plant Biol 17: 14
- 244. Zarei B, Naderi A, Jalal Kamali MR, Lack S, Modhej A. 2013. Determination of physiological traits related to terminal drought and heat stress tolerance in spring wheat genotypes. Intl J Agri Crop Sci 5: 2511-2520.
- 245. Zhang B, Liu W, Chang SX, Anyia AO. 2010. Water-deficit and high temperature affected water use efficiency and arabinoxylan concentration in spring wheat. J Cereal Sci 52: 263-269.
- 246. Zhang J, Zhang H, Li S, Li J, Yan L, Xia L. 2021a. Increasing yield potential through manipulating of an ARE1 ortholog related to nitrogen use efficiency in wheat by CRISPR/Cas9. J Integr Plant Biol 63: 1649-1663.
- 247. Zhang S, Zhang R, Gao J, Gu T, Song G, Li W, Li D, Li Y, Li G. 2019. Highly efficient and heritable targeted mutagenesis in wheat via the Agrobacterium tumefaciens-mediated CRISPR/Cas9 system. Int J Mol Sci 20: 4257
- 248. Zhang S, Zhang R, Gao J, Song G, Li J, Li W, Qi Y, Li Y, Li G. 2021b. CRISPR/Cas9-mediated genome editing for wheat grain quality improvement. Plant Biotechnol J 19: 1684-1686.
- 249. Zhang Z, Hua L, Gupta A, Tricoli D, Edwards KJ, Yang B, Li W. 2019. Development of an Agrobacterium-delivered CRISPR/Cas9 system for wheat genome editing. Plant Biotechnol J 17: 1623-1635.
- 250. Zhao H, Dai T, Jiang D, Cao W. 2008. Effects of high temperature on key enzymes involved in starch and protein formation in grains of two wheat cultivars. J Agron Crop Sci 194: 47-54.
- 251. Zhao H, Dai T, Jing Q, Jiang D, Cao W. 2007. Leaf senescence and grain filling affected by post-anthesis high temperatures in two different wheat cultivars. Plant Growth Regul 51: 149-158.
- 252. Zhao Hui ZH, Dai TingBo DT, Jing Qi JQ, Jiang Dong JD, Cao WeiXing CW, Lu Wei LW, Tian XiaoWei TX. 2006. Effects of high temperature during grain filling on key enzymes involved in starch synthesis in two wheat cultivars with different quality types. Acta Agron Sin 32: 423-429.
- 253. Zhao K, Tao Y, Liu M, Yang D, Zhu M, Ding J, Zhu X, Guo W, Zhou G, Li C. 2022. Does temporary heat stress or low temperature stress similarly affect yield, starch, and protein of winter wheat grain during grain filling? J Cereal Sci 103: 103408.
- 254. Zheng Y, Cai Z, Wang Z, Maruza TM, Zhang G. 2025. The genetics and breeding of heat stress tolerance in wheat: Advances and prospects. Plants 14: 148
- 255. Zhu Y, Lin Y, Fan Y, Wang Y, Li P, Xiong J, He Y, Cheng S, Ye X, Wang F, Goodrich J, Zhu JK, Wang K, Zhang CJ. 2023. CRISPR/Cas9-mediated restoration of Tamyb10 to create pre-harvest sprouting-resistant red wheat. Plant Biotechnol J 21: 665-667.
- 256. Zmani MM, Nabipour M, Meskarbashee M. 2018. Effects of heat stress after anthesis on PSⅡ photochemical efficiency and the antioxidant activity of wheat cultivars. J Agri Sci 24: 72-81.