적요
Salt stress is a major abiotic factor that limits wheat production worldwide. However, this threat is increasing significantly because soil salinity affects approximately 20% of the irrigated agricultural land globally, leading to significant yield losses by impairing plant growth and photosynthetic efficiency. This study aimed to identify single-nucleotide polymorphisms (SNPs) associated with salt tolerance in wheat core collections during the heading stage under saline stress conditions. Chlorophyll content, a physiological indicator of salt tolerance at heading, and soil electrical conductivity (EC) were measured in 609 accessions and a Salt Tolerance Index (STI) was subsequently constructed. Genome-wide association studies (GWAS) were performed using a 35 K SNP chip to identify significant marker-trait associations. Three models (MLM, FarmCPU, and BLINK) were employed for the GWAS, with FarmCPU and BLINK demonstrating superior power over the MLM in controlling false positives. GWAS results revealed four significant SNPs (AX-94929101, AX- 94615611, AX-94510535, and AX-94411611) located on chromosomes 3D, 5D, and 7D. AX-94510535 exhibited significant phenotypic differences based on SNP genotype, suggesting its potential as a marker for STI. Furthermore, the identified candidate genes, TraesCS3D02G218100, TraesCS5D02G059500, and TraesCS5D02G175000, were implicated in biological processes such as DNA replication, cell death, and photosynthesis.
서언
최근 세계적인 이상기후 현상과 다양한 환경오염 문제의 심화로 인해 밀 생산은 여러 형태의 환경 스트레스로부터 심각한 위협을 받고 있으며 염분으로 인하여 생장 장애, 생식 불량이 나타나고 밀 생산량이 6% 줄어들 것으로 예상되고 있다(
Asseng et al. 2015,
Turan et al. 2009). 특히, 토양 염류 집적에 의한 염 스트레스(salt stress)는 고온, 가뭄, 수분장해 등과 함께 전 세계적으로 문제가 되는 주요 환경 스트레스로 여겨지며, 농업 생산성에 중대한 영향을 미치는 요인으로 부각되고 있다(
Munns & Tester 2008). 실제로 전 세계의 관개 농경지 약 20%가 염류 집적의 영향을 받고 있으며, 염 스트레스가 작물에 미치는 부정적인 영향은 점점 뚜렷해지고 있다(
Munns & Gilliham 2015). 또한, 전 세계적으로 관개 농경지의 염해가 지속적으로 확대될 것으로 예상되기 때문에 염 스트레스에 대응하기 위한 농업 및 육종 연구의 중요성이 더욱 주목받고 있으며 염화된 토양에서의 안정적인 작물 생육과 대응하는 재배 기술 확보 및 저항성 작물 개발은 필수적인 연구 주제이다(
Flowers et al. 2010,
Qadir et al. 2008,
Rozema & Flowers 2008).
염 스트레스 조건에서 작물은 적응성을 높이기 위하여 다양한 생리적, 생화학적 반응을 일으킨다. 특히, 염 스트레스의 노출은 Na⁺ 이온의 과도한 축적을 유도하며 이로 인한 낮은 삼투포텐셜(osmotic potential)은 K⁺ 흡수를 억제 시켜서 Na⁺/K⁺ 비율의 불균형을 초래한다(
Zelm et al. 2020). 이러한 이온 불균형은 식물 뿌리에서부터 주요 세포 및 기관에 걸쳐 필수 이온의 수송과 항상성을 방해하고 세포 내 대사 및 생장 과정의 장애를 유발한다(
Bassil et al. 2019,
Yarra 2019). 또한 광합성 효율 저하, 막 구조 손상, 활성산소종(reactive oxygen species, ROS) 축적, 효소 활력 저하, 호르몬 신호전달 등의 식물의 필수 생리 대사 요소들의 변화가 나타난다(
Hasanuzzaman et al. 2014,
Hossain et al. 2018,
Wani et al. 2016). 이처럼 염 스트레스는 단순한 세포 내 대사 교란을 넘어 세포 기능과 다양한 생리적 과정에 광범위한 변화를 유발하며, 이러한 변화는 식물의 유전자 발현 패턴과 대사 경로 조절에도 반영된다(
Zelm et al. 2020). 결과적으로 염 스트레스에 적응하지 못하는 작물은 생장 저해, 발달 지연, 생식 장애 및 불임 등을 겪으며 이는 수확량 감소를 포함한 전 생애에 걸친 문제로 이어져 농업 생산성과 작물 품질에 중대한 영향을 미친다.
최근 다양한 작물에서 GBS (Genotyping-by-Sequencing) 기반 유전자형 데이터가 축적됨에 따라, 이를 활용한 전장유전체연관분석(Genome-Wide Association Study, GWAS)이 주요 형질의 유전적 분석 도구로 널리 활용되고 있다(
Javid et al. 2022,
Min et al. 2024,
Rabieyan et al. 2022). 특히, 유전체가 크고 복잡한 밀과 같은 거대배수성 작물은 환경 스트레스 내성 형질을 규명하기 위해서 연관불균형(Linkage Disequilibrium, LD)을 기반으로 하여 특정 형질과 연관된 유전자 또는 QTL (quantitative trait locus)의 위치를 높은 해상도로 탐색할 수 있는 GWAS연구가 적극적으로 활용되고 있는데 이는 기존의 QTL 분석보다 정밀한 유전자 탐색이 가능하다는 장점을 지닌다(
Kang et al. 2023,
Min et al. 2024,
Rabieyan et al. 2022). 또한, 이번 연구에 활용된 핵심집단은 전체 유전자원의 다양성을 최소한의 중복으로 대표할 수 있도록 설계된 자원 집단으로, GWAS 분석의 효율성을 크게 향상시킬 수 있으며 복잡성으로 인한 연구상의 제약을 극복하는 하나의 효과적인 전략으로 활용할 수 있다(
Balfourier et al. 2007,
Kang et al. 2023). 따라서, 밀의 내염성 유전자를 탐색하기 위해서는 정밀한 표현형 데이터를 유전체 데이터와 연관 분석을 수행함으로써 염 스트레스에 연관된 유의한 SNP 및 QTL을 발굴하고, 이를 기반으로 유용 대립유전자의 확보, 분자표지 개발 및 MAS (Marker-assisted selection) 등 내염성 품종 개발에 활용할 수 있는 실질적인 유전적 자원을 확보하는 것이 중요하다(
Devate et al. 2022).
본 연구에서는 한국형 밀 핵심집단을 활용하여 생식생장기에서 중요한 시기인 출수기에 NaCl 스트레스를 처리하여 SPAD value 및 토양 용존고형물 농도를 조사하였다. 이에 염 스트레스 저항성 계통을 탐색하고 GWAS 분석을 수행하여 연관된 SNP의 마커 활용 가능성과 후보 유전자군을 식별하고자 한다.
재료 및 방법
핵심집단 공시재료 및 생육환경
한국형 밀 핵심집단 609점을 농촌진흥청에서 분양 받아 공주대학교 연구 포장(충남 예산, (N): 36°66'80.02", (E): 126°85'99.944")에 파종하였다. 실험에 사용된 한국형 밀 핵심집단에는 국내 대표 장려 품종인 금강밀을 포함한 5개의 국내 품종도 포함되어 있다(
Min et al. 2021). 핵심집단에 파종된 포장은 각 계통마다 1 m간격으로 1 m에 약 35립 정도씩 2021년 10월 18일에 파종하였다. 내염성의 표현형 데이터 조사는 609점의 핵심집단 중 국내 생육환경에 생존한 567계통의 식물체를 절간신장기에 55.8 cm×19.6 cm×15.8 cm의 초화박스에 옮겨 심어 비가림 온실에서 출수를 진행하였다. 이식날짜는 국내 대표품종인 금강밀의 출수기 2일 전에 진행하였으며 염처리는 이식 후 1주일 뒤에 진행하였다.
한국형 밀 핵심집단의 염처리는 3일동안 200 mM의 NaCl 수용액 50 mL를 각 포트에 관수처리 방식으로 진행하였으며 3일간의 염처리 다음날에 엽록소를 3반복 측정하고 토양에서의 전기전도도도 측정하였다. 4일간의 처리(3일간의 염처리와 1일간의 측정)를 1유닛으로 정의하고 12일 동안 총 3유닛의 데이터를 확보하였다. 유닛에 적용된 계통 당 50 mL/일의 관수방법은 식물 생장을 위한 최소 수분으로서 농촌진흥청의 토양수분 장력 측정 시 사용되는 기준을 적용하였다(RDA 2012). 염 스트레스 처리구에서 각 계통의 지엽에서 3반복으로 SPAD-502 (KONICA MINOLTA, Tokyo, Japan)를 사용하여 엽록소 형광값을 측정하였으며 HMM-200 pro (Han young systems, Seoul, Korea) 토양수분 및 전기전도도계를 통해 전기전도도를 측정하였다.
핵심집단 출수기의 염 스트레스 지수(STI, Salt Tolerant Index)는 각 핵심집단 계통별 엽록소의 함량을 토양의 전기전도도에 토양상관계수(Correlation soil Electrical Conductivity: 640)을 곱한 토양의 총용존고형물농도로 나누어 계산하였다(
Mohamedin et al. 2006,
Rusydi 2018). 또한, 각 유닛별 핵심집단의 계통에 대한 STI를 구하여 GWAS에 활용할 표현형 데이터로 사용하였다.
패트리 디시에서 자란 10일 된 어린 잎을 채취하여 액화질소를 활용하여 고운 가루로 갈아준 후 Genomic DNA Prep Kit (Biofact, Daejeon, Korea)를 활용하여 Genomic DNA를 확보하였으며 확보된 DNA 농도는 20 ng/ul으로 조절하여 분석할 때까지 -20℃에 보관하였다. 유전형 데이터는 한국형 밀 핵심집단 전체 609점 중 DNA QC (DQC of 0.82, QC call rate 97%)조건과 국내 재배조건을 기준으로 하여 정상적 생육이 가능한 계통으로 선발하여 최종 567점을 이용하여 분석하였다. 밀 핵심집단 내 567점의 DNA를 35,143개의 SNP마커가 포함된 Axiom회사의 상용 SNP array인 BreedWheat 35K SNPchip (Thermo Fisher Scientific, MA, USA)을 이용하여 유전형 데이터를 확보하였고 Axiom Analysis Suite 5.2 소프트웨어(Thermo Fisher Scientific, MA, USA)로 분석하였다(
https://www.cerealsdb.uk.net/cerealgenomics/cerealsdb/axiom_download.php). Tassel v5.2.57 프로그램을 활용하여 분석된 SNP chip 데이터 중에서 minor allele frequency (MAF)가 5% 미만이면서 missing value가 10% 초과되는 SNP들을 적합하지 않은 SNP로 판단하여 제거하였으며, Beagle v5.2를 활용하여 imputation을 수행하여 NN 유전형을 제거하여 SNP chip 필터링을 진행하였다(
Bradbury et al. 2007,
Browning et al. 2018)
수집한 표현형 데이터인 염 스트레스 지수와 SNP chip data를 활용하여 R 프로그램에서의 GAPIT version3 패키지를 활용하여 각 유닛에 대한 GWAS를 수행하였다. GWAS 분석에 사용된 모델은 mixed linear model (MLM), Fixed and random model circulating probability unification (FarmCPU), Bayesian-information and linkage-disequilibrium iteratively nested keyway (BLINK) 3가지 모델로 수행하였으며 각 모델별로 도출된 결과를 서로 비교분석 하였다.
염 스트레스 연관 SNP 발굴 및 후보 유전자 발굴
GWAS 결과로 선발된 SNP들에 대하여 표현형데이터를 대입하여 상,하위 5계통들의 유전형과 같이 조사하여 SNP의 차이로 염 스트레스 지수를 판별할 수 있는 SNP 마커의 가능성을 판별하였다. GAPIT version3의 cut off 기준을 토대로 Bonferroni correction (0.05)를 차용하여 실제 GWAS에 사용된 SNP 마커의
p-value값을 log로 치환하여 최종적으로 LOT (Limit of Threshold) 5.65 이상의 SNP를 선발하였다. 또한, 선발된 SNP에 대해 567계통의 haplotype 분석을 통하여, 마커로서의 활용 가능성을 검정하였다. 후보유전자군의 경우 IWGSC v1.0 genome sequence를 참조하여 SNP를 기준으로 ±250 kb 탐색하였다. 선발 SNP 기준 ±250 kb에서 후보유전자를 선발한 이유는 선행연구에서 밀 염색체에서 LD값이 최소 74.7 kb, 최대 393 kb가 보고되었기에 대략적인 평균값인 250 kb에서 후보유전자를 추정하였다(
Kang et al. 2023,
Li et al. 2019,
Zhao et al. 2020,
Zhu et al. 2021).
결과 및 고찰
엽록소 및 토양의 염류농도 분석
공주대 연구 포장에서 생육 중인 밀 핵심집단 계통 중에서 국내 재배조건에서 정상 생육하는 567 계통에 대하여 표현형 데이터 및 유전형 데이터를 확보하였다. 표현형 데이터는 생식생장기인 출수기에 염처리와 무처리 집단에 대하여 엽록소와 전기전도도를 측정하여 분석하였다. 유닛1에서의 엽록소 최댓값은 51.46 mg/m
2, 최솟값은 12.4 mg/m
2로 유닛2에서는 최댓값은 55.63 mg/m
2, 최솟값은 4.2 mg/m
2로 유닛3에서는 최대 55.3 mg/m
2, 최소 0.7 mg/m
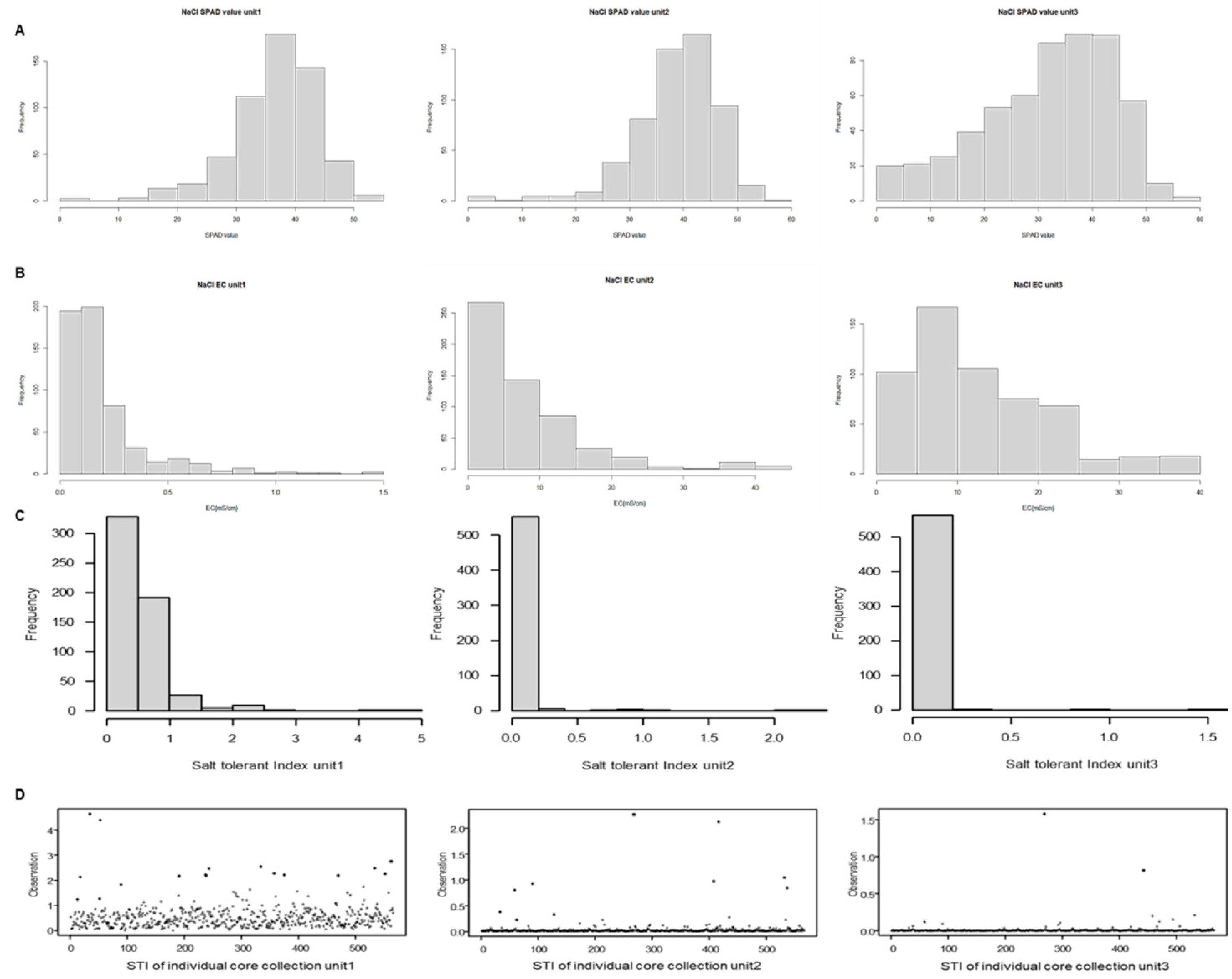
2로 나타났다. 염분이 누적된 유닛3에서는 엽록소의 최대값은 큰 차이가 없었으나 최소값에서는 크게 감소하는 것을 확인할 수 있었다. 전기전도도의 경우 최초로 측정 시에 모든 토양의 평균값이 0.064 mS/cm 였으나 시간이 경과함에 따라 유닛3에서의 토양의 전기전도도 평균값은 39.4 mS/cm로 나타났다. 이는 시간이 경과함에 따라 염 처리로 인하여 SPAD value가 감소하였으며, 토양에서는 NaCl 염이 누적되었기 때문으로 예상된다.
Rhee et al. (2007) 에 따르면 토양에서의 높은 염류 농도로 인해 작물에서의 광합성 속도가 감소하게 되는데 이는 기공이 닫히는 현상 뿐만 아니라 SPAD value의 감소도 주된 원인으로 꼽았다. 또한, SPAD value만으로 내염성 저항성을 판별하기 어려워 토양에 있는 용존고형물농도 대비 SPAD value인 STI를 제시하였으며 STI를 기준으로 조사한 567계통의 1%에 해당하는 상하위 5계통을 유닛별로 나타내었다(
Table 1). 출수기에서의 STI지수를 유닛별로 보았을 때, 유닛1에서는 최솟값 0.024, 최댓값 4.654로, 유닛2에서는 최솟값 0.0003, 최댓값 2.2701로, 유닛3에서는 최솟값 0.0001, 최댓값 1.579로 나타났다. 이는 유닛1에서 유닛3으로 염 스트레스가 누적됨에 따라 식물에서의 SPAD value는 염 스트레스를 받아 낮아지면서 토양에 집적되는 용존고형물농도는 증가하여 최솟값과 최댓값이 전부 줄어드는 경향을 볼 수 있다. 567계통에 대하여 STI지수를 유닛별 전체 분포도로 나타내었을 때, 유닛1보다 유닛2, 3에서 STI 값이 좌측으로 치우쳐져 있는 것으로 관찰되었다(
Fig. 1). 이는 염해 스트레스가 누적될수록 소수의 내염성 계통이 SPAD value가 높거나 토양 내 염류농도를 감소시킨 것으로 추론할 수 있다(
Fig. 1). 상위 계통 중에 cc1687과 cc1066 계통은 높은 STI값을 공통적으로 띄고 있어 내염성 자원 후보로 선발하였으며, 반면에 cc267의 경우에는 공통적으로 낮은 STI값을 가지고 있어 염 스트레스에 민감한 계통임을 알 수 있다.
핵심집단 중 567계통에 대하여 전체 35,143개의 SNP마커 중에 SNP의 위치가 명확하게 밝혀지지 않은 마커들과 MAF (Minor allele frequency)값 0.05 미만을 필터링하여 결과적으로 Axiom사의 BreedWheat 35K SNP chip에서 22,775개의 SNP들을 확보하였다. 이러한 SNP들은 모든 염색체에 분산되어 있는데, A 게놈에는 7,065개의 SNP 마커가 포함되어 있고, B 게놈에는 8,480개의 마커가 있으며, D 게놈에는 7,230개의 마커가 존재하였다. 밀은 A, B, D 게놈으로 구성되어 있는데 22,775개의 SNP 마커 분포를 살펴보았을 때, A 게놈에서 31.1%, B 게놈에서 37.2%, D 게놈에서 31.7%로 이루어져 있으며, B 게놈이 다른 게놈보다 약간 더 많은 SNP 마커를 포함하고 있다는 것으로 나타났다.
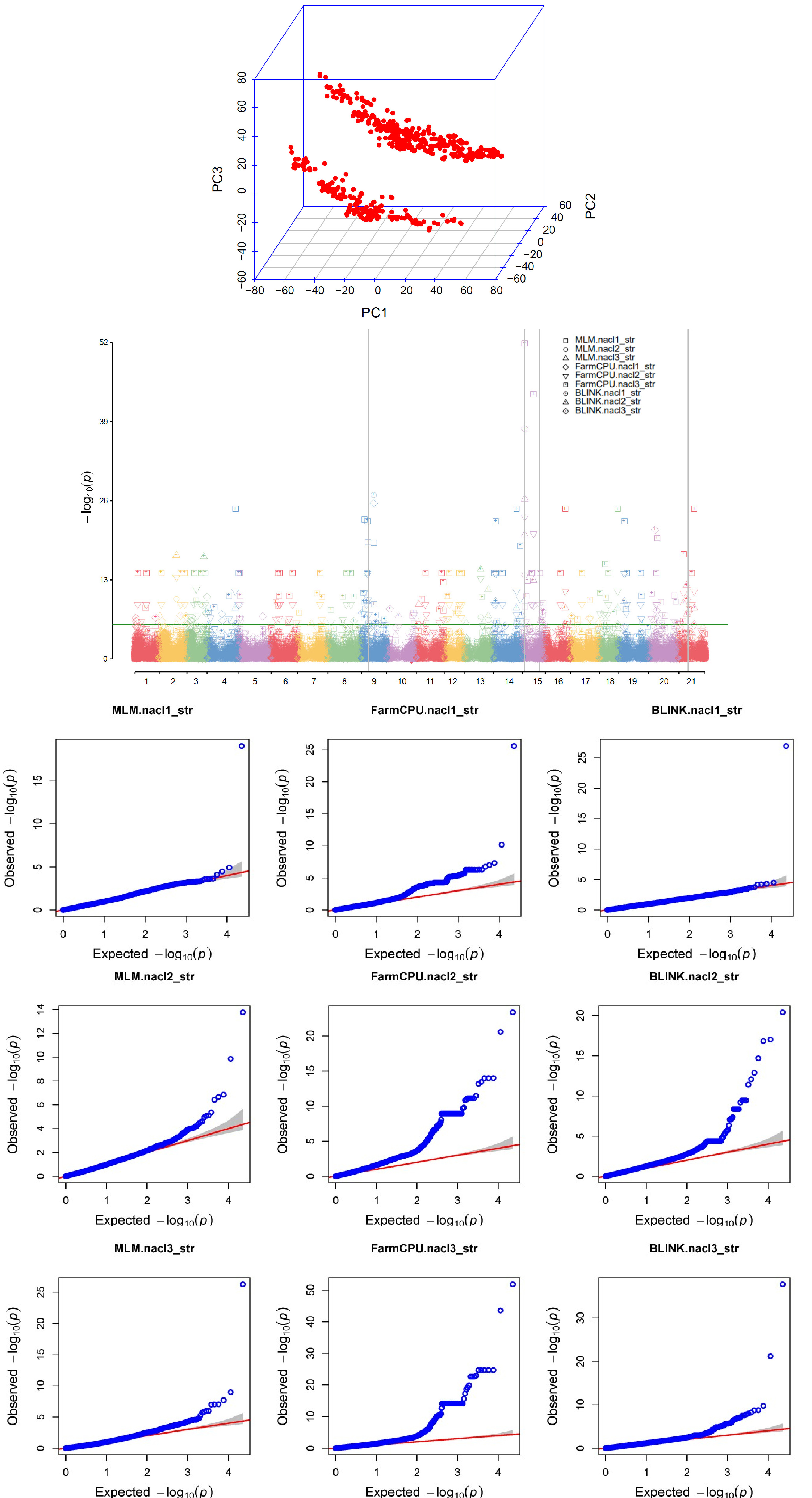
출수기의 STI에 대한 GWAS 분석은 MLM, FarmCPU, BLINK의 3가지 모델을 이용하였으며 분석을 통하여 발굴된 SNP 연관 지역에 대하여 유전자 후보군을 탐색하였다(
Fig. 2). MLM의 경우에는 고정효과와 임의효과를 모두 포함한 혼합선형 모델이며, FarmCPU의 경우에는 친족관계 행렬 재구성을 활용하여 통계적 검정력이 향상되었으며, BLINK의 경우에는 LD정보를 활용하는 특징이 있다(
Cebeci et al. 2023).
GWAS결과 LOT 5.65이상의 값을 가지는 SNP들은 다수 발굴되었으나, 재현성 있는 SNP 발굴을 위하여 GWAS 모델별로 선발된 SNP중 공통으로 발견된 SNP들을 선발하였다. 또한 염처리의 누적된 효과를 확인하기 위하여 각 유닛별로 공통으로 발굴된 SNP를 선발하였다. AX-94411611의 경우 유닛 2와 유닛 3에서 오직 BLINK 모델에서 발견되어 총 2번 탐색되었으며, AX-94929101의 경우 MLM 모델에서는 탐색되지 않았으나, 유닛 2, 유닛 3에서의 FramCPU, BLINK 모델에서 탐색되어 총 4번 공통적으로 나타났으며, AX-945010535는 유닛 1에서의 모든 모델에서 탐색되어 3번, AX-94615611은 유닛 2, 유닛3에서의 모든 GWAS 모델에서 탐색되어 총 6번 탐색되었다(
Table 2).
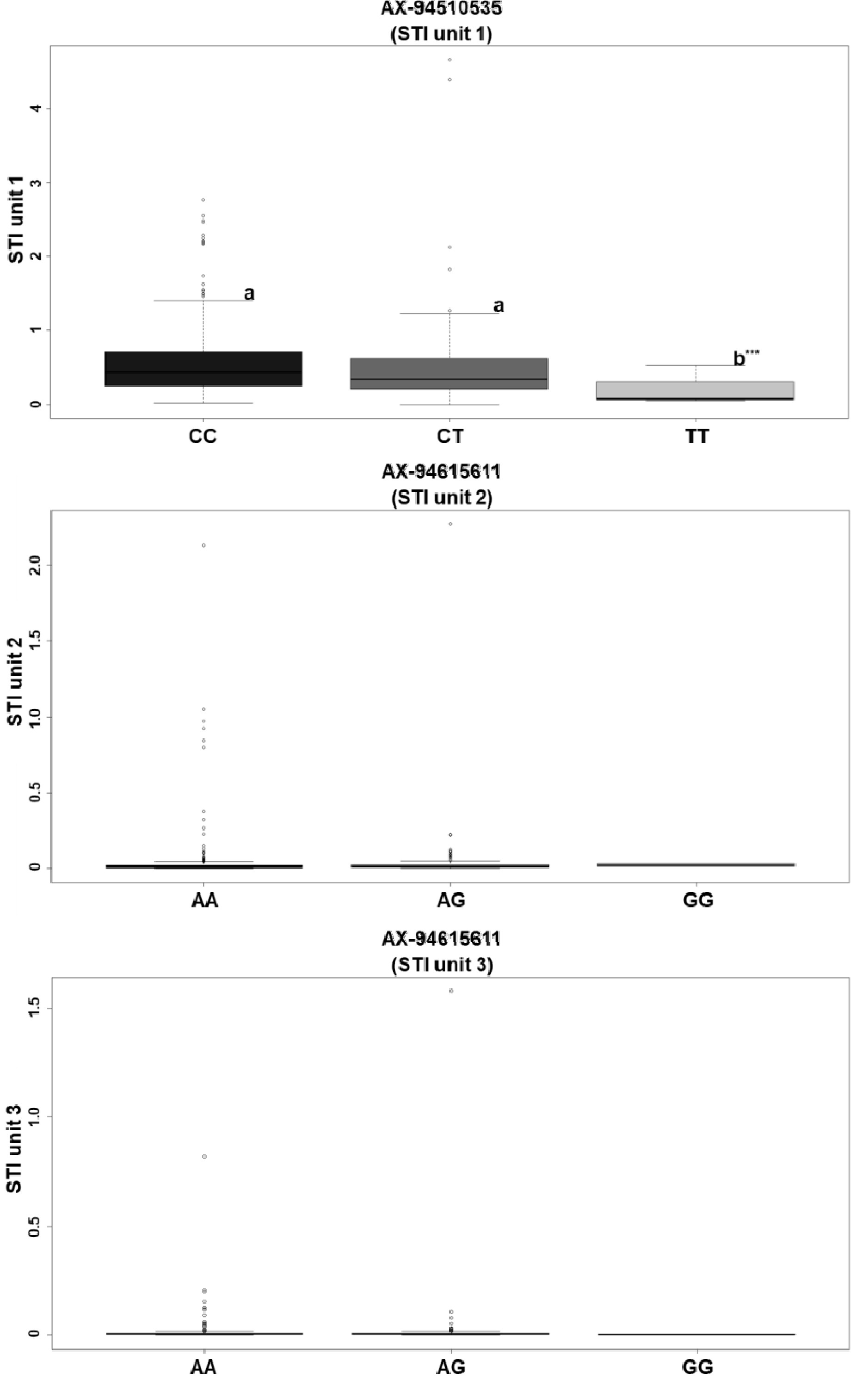
결과적으로 AX-94411611, AX-94615611, AX-94929101, AX-94510535의 SNP가 내염성 후보 SNP로 탐색되었으며, 각각 3D, 5D, 7D 염색체에서 유의성을 보이는 연관분석 결과가 나타났다. 선발된 SNP들에 마커로서의 가능성을 판별하기 위하여 던컨의 다중검정(Duncan's multiple tsest) (alpha=0.05)을 수행하였다. 분석을 수행한 결과 AX-94411611의 경우에 각각 ANOVA분석 결과 Pr값이 0.958 (유닛 2) 0.882 (유닛 3)으로, AX-94615611의 경우에는 Pr값이 0.475 (유닛 2), 0.460 (유닛 3)으로 나타나 마커로서의 가능성이 나타나지 않았다. 특히 AX-94929101은 핵심집단 내에서 모든 계통이 표현형 데이터와 관계 없이 유전형이 이형접합인 AG만 나타났으므로 마커로서의 활용가능성이 없으며, 따라서 다중비교를 수행하지 못하였다. 반면 AX-94510535의 경우에는 마커 활용 가능성이 나타났으며, 유전형 CT와 CC는 a그룹으로 상위계통을 대표하였으며, TT는 b그룹으로 하위계통에서 주로 발견되었다(Pr=0.0000783) (
Fig. 3,
Table 3). 한 개의 SNP로 양적형질인 내염성을 판단하기에는 무리가 있으나 발굴한 유전자 후보군들의 염기서열을 참조하여 염 스트레스에 영향을 미치는 SNP를 탐색할 수 있을 것으로 사료된다.
출수기에서 STI 표현형 데이터에서 탐색된 SNP들 중에서 MLM, FarmCPU, BLINK에서 공통적으로 발견된 SNP인 3개의 SNP들 주변으로 ±250 kb를 탐색한 결과, uncharacterized protein을 제외하고 총 12개의 유전자 후보군을 확인하였다(
Table 4). 또한, 단형성 마커인 AX-94929101은 마커로서의 가능성은 없지만
Skovbjerg et al. (2025)에 기반하여 유전자 후보군을 탐색하게 되었다. AX-94929101은 출수시기의 유닛 1 표현형데이터에 의해서 발견되었는데 유전자 내에 존재하는 SNP로서, 염기서열이 G또는 A가 치환될 경우, 아미노산 서열이 바뀌는 missense 돌연변이를 유발하게 된다. missense SNP인 AX-94929101가 속한 TraesCS3D02G218100 (AAA+ ATPase domain-containing protein)에서 AAA+단백질은 ATP를 에너지원으로 활용하여 기능적으로나 구조적으로 다양하며 이들은 DNA 복제, 단백질 분해, 신호 전달 및 유전자 발현 조절 등과 같은 과정에 관여한다(
Erzberger & Berger 2006,
Miller & Enemarkal 2016). 또한, 주변에서 발견된 TraesCS3D02G218400 (C3HC-type domain-containing protein)은 CCCH zing-finger단백질이 함유하고 있는 모티프로 애기장대에서 염분 스트레스 조건에서 생육 실험을 진행하여
Ghc3h20 유전자 형질전환 애기장대와 일반 애기장대를 비교하여 염 스트레스 내성을 향상시키기 위한 ABA 신호 경로에 관여하는 것으로 밝혀졌다(
Zhang et al. 2023). 이릍 통하여 내염성과 연관되어 있는 유전자로 예상된다. AX-94510535의 경우 전사체의 엑손 부분에 속하지 않고 3' UTR영역에 속하는데 이 SNP가 포함되어 있는 TraesCS5D02G059500 (Zinc finger LSD1-type domain-containing protein)에서의 Zinc finger LSD1-type은 애기장대에서 세포의 사멸과 병저항성에 연관되어 있다고 알려져 있으며,
Guo et al. (2013)의 연구에서는 밀 줄기녹병을 연구하면서
TaLSD1 을 특정하였다. 곰팡이와 같은 병원체의 감염에 의해 세포 사멸 시에 음성 조절자로 작용하는 것으로 보아 식물의 스트레스 반응에 중요한 역할을 하여 위의 유전자 후보군도 내염성과 연관이 있을 것으로 사료된다(
Guo et al.2013). AX-94411611의 경우에는 엑손 부분이 아닌 인트론부분에 속해 있으며 이 SNP가 속해 있는 TraesCS7D02G233900는 아직 아무런 기능이 밝혀지지 않은 Uncharacterized protein로 나타났다. 가장 많은 GWAS모델에서 발견된 AX-94615611도 AX-94929101과 같이 엑손에서의 염기서열 치환이 아미노산에 영향을 미치는 missense로 나타났다. 이 SNP가 속한 TraesCS5D02G175000 (Phosphoglycolate phosphatase)에서의 포스포글리콜산 포스파타아제는 광호흡 과정에서 2-Phosphoglycolate를 글리콜산으로 변환시키는 기작에 관여하며, 광합성 과정에서 캘빈 회로의 효율성을 유지하게 함으로서, 손상된 원형질체에서의 세포의 자가분열 안정성을 도모한다(
Ni et al. 2022). 이를 통하여 염 스트레스를 받아 SPAD value가 감소함에 따라 세포적 차원에서의 손실을 막을 수 있게 도와주는 효소로 내염성과 연관된 유전자 후보로 사료된다. 또한, AX-94615611 주변에서 탐색된 TraesCS5D02G175100 (GBF-interacting protein 1 N-terminal domain-containing protein)에서의 GBF는 bZIP 단백질 계열로 식물의 환경적 변이에 연관되어 있는 것으로 알려져 있으며, 내냉성, 내염성, 건조, 광, abscisic acid 등에 이르는 광범위한 반응에 대응하는 핵심 요소로 알려져 있으므로 내염성 기작에 관여 할 것으로 예상된다(
Menkens et al. 1995, Shinozaki & Yamaguchi-shinozaki 1997).
본 연구는 한국밀 핵심집단의 출수기에 염해 스트레스를 처리하여 SPAD value와 토양전기전도도를 측정하였으며, 토양의 총용존고형물 농도(염류농도)와 SPAD value를 활용하여 STI지수를 3개의 유닛에서 조사하여 집단 내 내염성 우수자원 선발과 연관된 SNP를 탐색하였다. 표현형 분석을 통하여 핵심집단 내에서 내염성 우수자원으로 2개의 계통(cc1687과 cc1066)을 선발하였으며 GWAS 연관 분석을 통하여 AX-94510535를 선발하였으며 선발된 SNP는 출수기의 내염성 마커로 개발될 가능성이 있는 것으로 확인되었다. 또한 발굴된 내염성 후보 유전자는 추가적인 분석을 통하여 내염성 밀 육종과 더불어 추후 간척지와 같은 염분이 높은 토양에서도 생육이 가능하게 되어 수확량 향상에 활용될 수 있을 것이다.
적요
본 연구는 밀의 이삭이 나오는 출수기에 염해 스트레스를 처리하고 유닛별 SPAD value 및 토양전기전도도를 활용한 STI지수를 기반으로 내염성 계통을 선발하고 GWAS 분석을 통해 염해 스트레스와 연관된 SNP 및 유용유전자 후보군을 탐색하였다.
1. 밀의 핵심집단내에 계통번호 cc1687과 cc1066 계통은 유닛2와 유닛3의 STI 지수에서 공통적으로 높은 값을 띄어 내염성 우수자원 후보로 선발하였음.
2. 전장유전체 연관분석(GWAS) 결과 3D, 5D, 7D에서 유의성이 있는 SNP를 발굴하였으며 발굴된 SNP는 AX- 94929101, AX-94510535, AX-94411611, AX-AX-94615611 이며 각각 엑손, 3'UTR, 인트론영역에서 존재하였음.
3. 탐색된 SNP를 중심으로 250kbp를 탐색한 결과 TraesCS3D02G218100(AAA+ ATPase domain-containing protein), TraesCS3D02G218400 (C3HC-type domain-containing protein), TraesCS5D02G059500 (Zinc finger LSD1- type domain-containing protein), TraesCS7D02G233900 (Uncharacterized protein), TraesCS5D02G175000 (Phosphoglycolate phosphatase), TraesCS5D02G175100 (GBF-interacting protein 1 N-terminal domain-containing protein)를 내염성 연관 candidate gene으로 선발하였음.
4. 선발 SNP의 핵심집단 내 하플로타입 분석을 통해 AX-94510535가 유전자형에 따른 표현형이 유의성이 나타났으므로 추후 STI 지수를 기준으로 내염성 계통을 선발할 마커로 활용이 가능할 것으로 사료됨.
사사
본 연구는 교육부 산하 한국연구재단의 지역대학우수과학자지원사업(NRF-2020R1I1A3069901)의 지원으로 수행되었습니다.
Fig. 1The each unit SPAD value(A), electrical conductivity (EC) (B), Salt tolerant index (STI) (C,D). STI distribution across the three units reveals that in Unit1, the values range from 0 to 1. In contrast, Units2 and 3 exhibit STI values predominantly between 0 and 0.5, with a small number of salt-tolerant candidate accessions displaying considerably higher values.
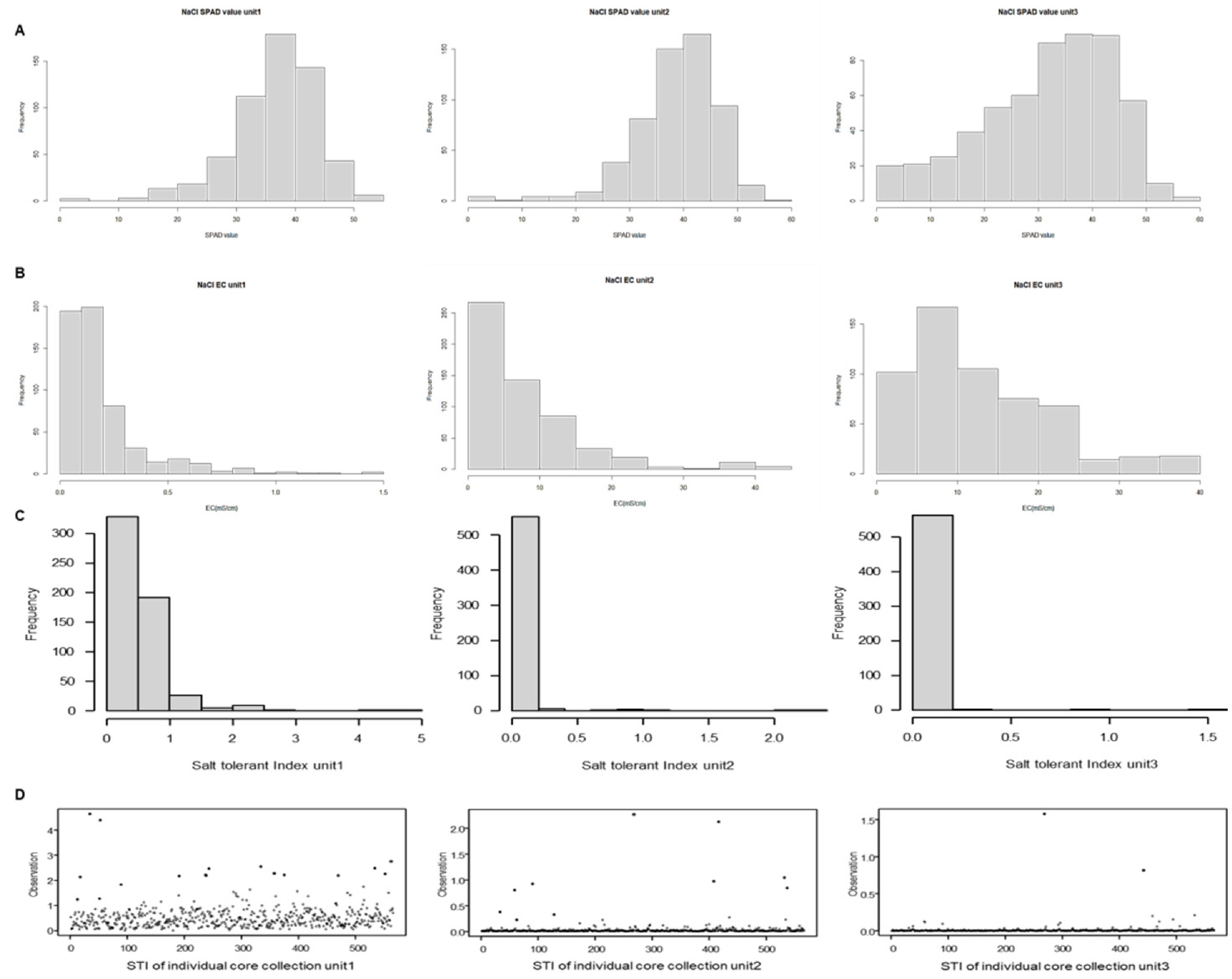
Fig. 2GWAS analysis of wheat core collection under saslt stress. PCA plot (A), Manhatta plot (B) for three models (MLM, FarmCPU, BLINK) and QQplot (C) to STI phenotype data GWAS result. SNP code “AX-94411611, AX-94615611, AX-94929101, and AX-94510535” was commonly found in the MLM model, FarmCPU model and the BLINK model. The SNPs connected by the gray lines in the Manhattan plot are common SNPs and selected SNPs. For qqplot, three models are included as results from STI all unit.
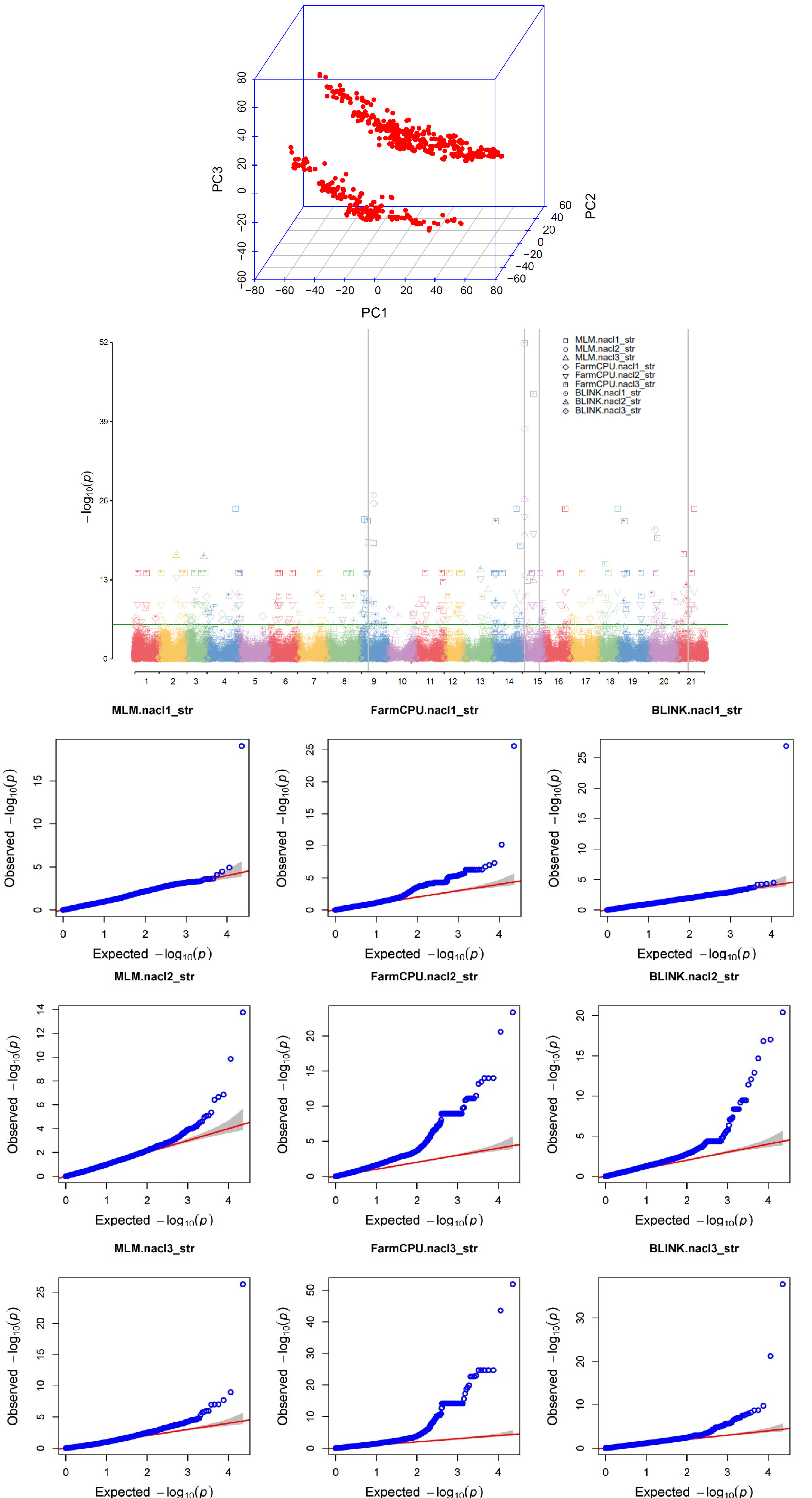
Fig. 3Box plot analysis for AX-94510535 and AX-94515611. Means with different lowercase letters in the figure are significantly different (p<0.05) to ANOVA and Duncan multiple test. The relevance of ax-94515611, ax-94411611, and ax-94929101 as markers has not been established.
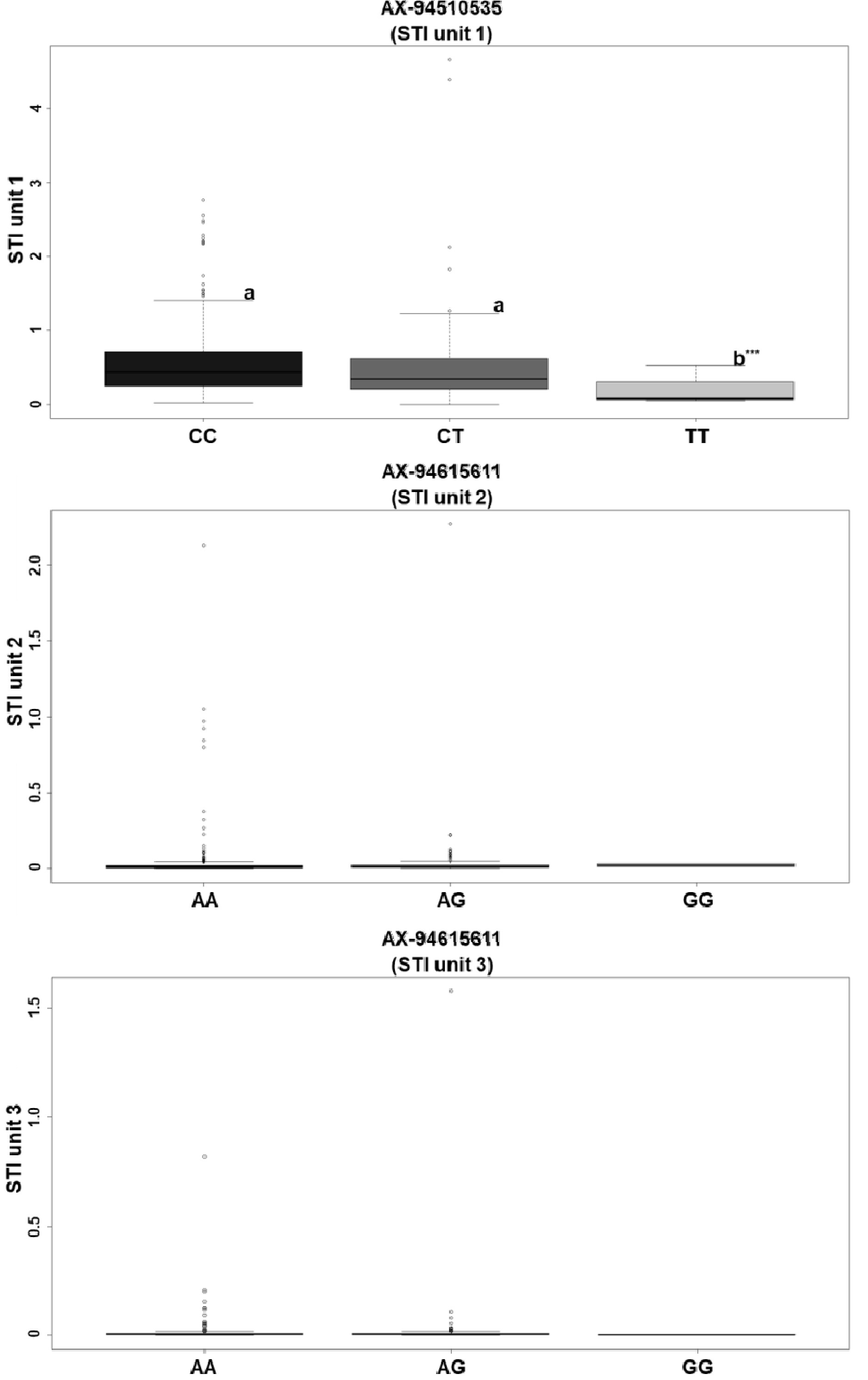
Table 1Summary of STI value of top 5 and bottom 5 accessions in heading stage under salt stress in each unit.
Table 1
|
STI-unit1 |
|
STI-unit2 |
|
STI-unit3 |
|
|
|
Core collection
No. |
Value |
Core collection
No. |
Value |
Core collection
No. |
Value |
|
Top5 |
cc845 |
4.654 |
|
cc1687 |
2.27 |
|
cc1687 |
1.57 |
|
cc621 |
4.392 |
|
cc1460 |
2.12 |
|
cc1526 |
0.82 |
|
cc1606 |
2.758 |
|
cc1066 |
1.04 |
|
cc1066 |
0.21 |
|
cc1213 |
2.547 |
|
cc594 |
0.97 |
|
cc901 |
0.20 |
|
cc1102 |
2.486 |
|
cc24 |
0.92 |
|
cc1824 |
0.15 |
|
Bottom5 |
cc145 |
0.024 |
|
cc267 |
0.0003 |
|
cc276 |
N.Dz
|
|
cc181 |
0.042 |
|
cc216 |
0.0007 |
|
cc676 |
N.D |
|
cc930 |
0.043 |
|
cc836 |
0.0011 |
|
cc678 |
N.D |
|
cc113 |
0.048 |
|
cc922 |
0.0011 |
|
cc267 |
N.D |
|
cc115 |
0.048 |
|
cc406 |
0.0012 |
|
cc163 |
0.0001 |
Table 2Significant SNPs discovered via multi models using the STI value data in GWAS results.
Table 2
|
SNP code |
Chromosome |
Positionz
|
Alleley
|
Model |
Trait |
Repetition |
|
AX-94929101 |
3D |
297,464,485 |
A/G |
FarmCPU, BLINK |
STI unit2, STI unit3 |
4 |
|
AX-94615611 |
5D |
274,571,559 |
T/C |
MLM, FarmCPU, BLINK |
STI unit2, STI unit3 |
6 |
|
AX-94510535 |
5D |
56,449,445 |
T/C |
MLM, FarmCPU, BLINK |
STI unit1 |
3 |
|
AX-94411611 |
7D |
195,633,401 |
C/T |
BLINK |
STI unit2, STI unit3 |
2 |
Table 3Number of haplotype variation in AX-94510535 related STI in the wheat core collection.
Table 3
|
AX-94510535_STI unit1 |
Mean |
Std. Deviation |
Std.Error |
No. of Accession |
|
|
Reference |
C |
|
Hap1 |
C |
0.550 |
0.444 |
0.019 |
382 |
|
Hap2 |
C/T |
0.564 |
0.123 |
0.058 |
4 |
|
Hap3 |
T |
0.420 |
0.287 |
0.029 |
178 |
Table 4Investigation of candidate genes in the range of 250 kbp upper and lower sequences of each selected SNP.
Table 4
|
SNP code |
Gene ID. |
Protein name |
Gene position on chromosome |
AX-94929101
(missense) |
TraesCS3D02G218000 |
DNL-type domain-containing protein |
297,465,771-297,467,881 |
|
TraesCS3D02G218100z
|
AAA+ ATPase domain-containing protein |
297,463,657-297,465,738 |
|
TraesCS3D02G218200 |
Protein GAMETE EXPRESSED 3 |
297,491,530-297,494,800 |
|
TraesCS3D02G218300 |
Kinesin-like protein |
297,495,032-297,499,090 |
|
TraesCS3D02G218400 |
C3HC-type domain-containing protein |
297,594,170-297,599,022 |
AX-94510535
(3’prime UTR) |
TraesCS5D02G059500z
|
Zinc finger LSD1-type domain-containing protein |
56,445,757-56,449,637 |
|
TraesCS5D02G059600 |
CASP-like protein |
56,450,701-56,452,848 |
|
TraesCS5D02G059800 |
Methyltransferase domain-containing protein |
56,463,207-56,470,737 |
AX-94411611
(Intron) |
TraesCS7D02G233600 |
Zinc-ribbon 15 domain-containing protein |
195,395,797-195,396,039 |
|
TraesCS7D02G233700 |
NmrA-like domain-containing protein |
195,615,425-195,621,621 |
|
TraesCS7D02G233900z
|
Uncharacterized protein |
195,629,626-195,635,061 |
AX-94615611
(missense) |
TraesCS5D02G174900 |
F-box domain-containing protein |
274,569,491-274,572,692 |
|
TraesCS5D02G175000z
|
Phosphoglycolate phosphatase, EC:3.1.3.18 |
274,574,126-274,580,148 |
|
TraesCS5D02G175100 |
GBF-interacting protein 1 N-terminal domain-containing protein |
274,764,237-274,775,158 |
|
TraesCS5D02G175200 |
F-box domain-containing protein |
274,775,998-274,778,515 |
References
- 1. Ali N, Rahman UI, Badakshi F, Tariq MJ, Mujeeb-Kazi A. In: Munir O, Alvina G. 2020. Ensuring sustainable food security : Exploiting alien genetic diversity in wheat breeding for adaptation to emerging stresses. (Eds). Climate change and food security with emphasis on wheat. Academic Press, Massachusetts, United States: pp. 31-42.
- 2. Ashraf M, Harris PJC. 2013. Photosynthesis under stressful environments: An overview. Photosynthetica 51: 163-190.
- 3. Asseng S, Ewert F, Martre P, Rötter RP, Lobell DB, Cammarano D, Kimball BA, Ottman MJ, Wall GW, White JW, Reynolds MP, Alderman PD, Prasad PVV, Aggarwal PK, Anothai J, Basso B, Biernath C, Challinor Andrew J, De Sanctis G, Doltra J, Fereres E, Garcia-Vila M, Gayler S, Hoogenboom G, Hunt LA, Izaurralde RC, Jabloun M, Jones CD, Kersebaum KC, Koehler A-K, Müller C, Naresh Kumar S, Nendel C, O'Leary G, Olesen JE, Palosuo T, Priesack E, Eyshi Rezaei E, Ruane AC, Semenov MA, Shcherbak I, Stöckle C, Stratonovitch P, Streck T, Supit I, Tao F, Thorburn PJ, Waha K, Wang E, Wallach D, Wolf J, Zhao Z, Zhu Y. 2015. Rising temperatures reduce global wheat production. Nature Climate Change 5: 143-147.
- 4. Balfourier F, Roussel V, Strelchenko P, Exbrayat-Vinson F, Sourdille P, Boutet G, Charmet G. 2007. A worldwide bread wheat core collection arrayed in a 384-well plate. Theor Appl Genet 114: 1265-1275.
- 5. Bassil E, Zhang S, Gong H, Tajima H, Blumwald E. 2019. Cation Specificity of Vacuolar NHX-Type Cation/H+ Antiporters. Plant Physiol 179: 616-629.
- 6. Bradbury PJ, Zhang Z, Kroon DE, Casstevens TM, Ramdoss Y, Buckler ES. 2007. TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics 23: 2633-2635.
- 7. Browning BL, Zhou Y, Browning SR. 2018. A one-penny imputed genome from next-generation reference panels. Am J Hum Genet 103: 338-348.
- 8. Cebeci Z, Bayraktar M, Gökçe G. 2023. Comparison of the statistical methods for genome-wide association studies on simulated quantitative traits of domesticated goats (Capra hircus L.). Small Ruminant Res 227: 107053.
- 9. Devate NB, Krishna H, Parmeshwarappa SKV, Manjunath KK, Chauhan D, Singh S, Singh JB, Kumar M, Patil R, Khan H, Jain N, Singh GP, Singh PK. 2022. Genome-wide association mapping for component traits of drought and heat tolerance in wheat. Front Plant Sci 13: 943033.
- 10. Erzberger JP, Berger JM. 2006. Evolutionary relationships and structural mechanisms of AAA+ proteins. Annu Rev Biophys Biomol Struct 35: 93-114.
- 11. Flowers TJ, Galal HK, Bromham L. 2010. Evolution of halophytes: multiple origins of salt tolerance in land plants. Funct Plant Biol 37: 604-612.
- 12. Guo J, Bai P, Yang Q, Liu F, Wang X, Huang L, Kang Z. 2013. Wheat zinc finger protein TaLSD1, a negative regulator of programmed cell death, is involved in wheat resistance against stripe rust fungus. Plant Physiol Biochem 71: 164-172.
- 13. Hasanuzzaman M, Alam MM, Rahman A, Hasanuzzaman M, Nahar K, Fujita M. 2014. Exogenous Proline and Glycine Betaine Mediated Upregulation of Antioxidant Defense and Glyoxalase Systems Provides Better Protection against Salt-Induced Oxidative Stress in Two Rice (Oryza sativa L. ) Varieties. Biomed Res Int,
- 14. Hossain MA, Li ZG, Hoque TS. 2018. Heat or cold priming-induced cross-tolerance to abiotic stresses in plants: key regulators and possible mechanisms. Protoplasma 255: 399-412.
- 15. Javid S, Bihamta MR, Omidi M, Mohammadi M, Naghavi MR, Mardi M. 2022. Genome-Wide Association Study (GWAS) and genome prediction of seedling salt tolerance in bread wheat (Triticum aestivum L.). BMC Plant Biol 22: 581
- 16. Kang Y, Choi C, Kim JY, Min KD, Kim C. 2023. Optimizing genomic selection of agricultural traits using K-wheat core collection. Front Plant Sci 14: 1112297.
- 17. Kumar S, Li G, Yang J, Huang X, Ji Q, Liu Z, Ke W, Hou H. 2021. Effect of Salt Stress on Growth, Physiological Parameters, and Ionic Concentration of Water Dropwort (Oenanthe javanica) Cultivars. Front Plant Sci 12: 660409.
- 18. Li F, Wen W, Liu J, Zhang Y, Cao S, He Z, Rasheed A, Jin H, Zhang C, Yan J, Zhang P, Wang J, Xia X. 2019. Genetic architecture of grain yield in bread wheat based on genome-wide association studies. BMC Plant Biol 19: 168
- 19. Mehta P, Jajoo A, Mathur S, Bharti S. 2010. Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem II in wheat leaves. Plant Physiol Biochem 4: 16-20.
- 20. Menkens AE, Schindler U, Cashmore AR. 1995. The G-box: a ubiquitous regulatory DNA element in plants bound by the GBF family of bZIP proteins. Trends Biochem 20: 506-10.
- 21. Miller JM, Enemark EJ. 2016. Fundamental Characteristics of AAA+ Protein Family Structure and Function. Archaea 14: 9294307.
- 22. Min KD, Cho JW, Kim JY. 2024. Genome-Wide Association Studies for Drought Tolerance in Wheat Core Collection by Chlorophyll Measurement during Tillering and Ripening Stages. Korean J Crop Sci 69: 294-306.
- 23. Min KD, Kang YN, Kim CS, Choi CH, Kim JY. 2021. Genetic diversity and population structure of Korean common wheat (Triticum aestivum). Korean J Breed Sci 53: 277-289.
- 24. Mohamedin A, Abd El-Kader A, Badran N. 2006. Response of Sunflower (Helianthus annuus L.) to Plants Salt Stress under Different Water Table Depths. J Appl Sci Res 2: 1175-1184.
- 25. Munns R, Gilliham M. 2015. Salinity tolerance of crops - what is the cost? New Phytol 208: 668-673.
- 26. Munns R, Tester M. 2008. Mechanisms of salinity tolerance. Annu Rev Plant Biol 59: 651-681.
- 27. Ni J, Li Y, Xiang Y, Yang X, Jia L, Yue J, Wang H. 2022. Autophagic degradation of the chloroplastic 2-phosphoglycolate phosphatase TaPGLP1 in wheat. Plant Cell Rep 41: 473-487.
- 28. Qadir M, Tubeileh A, Akhtar J, Larbi A, Minhas PS, Khan MA. 2008. Productivity enhancement of salt-affected environments through crop diversification. Land Degrad Degrad Dev 19: 429-453.
- 29. Rabieyan E, Bihamta MR, Moghaddam ME, Mohammadi M, Naghavi MR, Mardi M. 2022. Genome-wide association mapping and genomic prediction for pre-harvest sprouting resistance, low α-amylase and seed color in Iranian bread wheat. BMC Plant Biol 22: 300
- 30. Rhee HC, Cho MW, Lee SY, Choi GL, Lee JH. 2007. Effect of salt concentration in soil on the growth, yield, photosynthetic rate, and mineratl uptake of tomato in protected vultivation. J Bio-environ Control 16: 328-332.
- 31. Rozema J, Flowers T. 2008. Crops for a salinized world. Science 322: 1478-1480.
- 32. Rural Development Administration (RDA).2012. Standard of research and analysis for agricultural technology.
- 33. Rusydi A. 2018. Correlation between conductivity and total dissolved solid in various type of water: A review. Conf Ser:. Earth Environ Sci 118: 012019.
- 34. Shinozaki K, Yamaguchi-Shinozaki K. 1997. Gene Expression and Signal Transduction in Water-Stress Response. Plant Physiol 115: 327-34.
- 35. Skovbjerg CK, Sarup P, Wahlström E, Jensen JD, Orabi J, Olesen L, Jensen J, Jahoor A, Ramstein G. 2025. Multi-population GWAS detects robust marker associations in a newly established six-rowed winter barley breeding program. Heredity 134: 33-48.
- 36. Turan MA, Elkarim AHA, Taban N, Taban S. 2009. Effect of salt stress on growth, stomatal resistance, proline and chlorophyll concentrations on maize plant. Afr J Agric Res 4: 893-897.
- 37. Wani SH, Kumar V, Shriram V, Sah SK. 2016. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J 4: 162-176.
- 38. Yarra R. 2019. The wheat NHX gene family: Potential role in improving salinity stress tolerance of plants. Plant Gene 18: 100178.
- 39. Zelm E, Zhang Y, Testerink C. 2020. Salt tolerance mechanisms of plant. Annu Rev Plant Biol 71: 403-433.
- 40. Zhang Q, Zhang J, Wei F, Fu X, Wei H, Lu J, Ma L, Wang H. 2023. The CCCH-type zinc-finger protein GhC3H20 enhances salt stress tolerance in Arabidopsis thaliana and cotton through ABA signal transduction pathway. Int J Mol Sci 24: 5057
- 41. Zhao G, Chang X, Wang D, Tao Z, Wang Y, Yang Y, Zhu Y. 2018. General situation and development of wheat production. Crops 34: 1-7.
- 42. Zhao Y, Li J, Zhao R, Xu K, Xiao Y, Zhang S, Tian J, Yang X. 2020. Genome-wide association study reveals the genetic basis of cold tolerance in wheat. Mol Breed 40: 36
- 43. Zhu T, Wang L, Rimbert H, Rodriguez JC, Deal KR, De Oliveira R, Choulet F, Keeble-Gagnère G, Tibbits J, Rogers J, Eversole K, Appels R, Gu YQ, Mascher M, Dvorak J, Luo MC. 2021. Optical maps refine the bread wheat Triticum aestivum cv. Chinese Spring genome assembly. Plant J 107: 303-314.